 |
|
|
| |
Biochemical Characterizations and drug sensitivity of the Effect of Mutations Selected in HIV-1 Integrase Gene Associated with Failure to Raltegravir (MK-0518)
|
| |
| |
Reported by Jules Levin
16th Intl HIV Drug Resistance Workshop, June 12-16, 2007, Barbados
Isabelle Malet1*,Olivier Delelis2, Marc-Antoine Valantin3,4, Brigitte Montes5, Cathia Soulie1, Marc Wirden1, Luba Tchertanova2, Gilles Peytavin6, Jacques Reynes3, Jean-François Mouscadet2, Christine Katlama3,4, Vincent Calvez1 and Anne-Genevieve Marcelin1
Laboratoire de Virologie, Hopital Pitie-Salpetriere, Assistance Publique-Hopitaux de Paris; EA 2387, Universite Pierre et Marie Curie (Paris 6), Paris,1 LBPA, CNRS, Ecole Normale Superieure de Cachan,2 INSERM U720, Universite Pierre et Marie Curie (Paris 6),3 Service des Maladies Infectieuses, Hopital Pitie-Salpetriere Paris4, Laboratoire de Virologie, Service des maladies infectieuses, Hopital Universitaire de Montpellier5, Service de Pharmacie Clinique, Hopital Bichat-Claude-Bernard, 46, rue Henri-Huchard, 75018 Paris6, France
The aim of this study was to describe the genetic changes ein the integrase gene of 4 heavily pretreated patients who received a salvage therapy including raltegravir and who subsequently developed virological failure under raltegravir therapy and to characterize the catalytic activities of wild-type and mutated recombinant integrases assayed in vitro and to test their sensibilities to raltegravir.
Author Conclusion: "The 4 studied patients showed the appearance of one or two mutations when they escaped from the MK-0518 treatment:G140S + Q148H, N155H, E92Q and E157Q, respectively. All mutations led to a resistance of the enzyme, thus confirming integrase as the primary target of the inhibitor in vivo. The fact that viral load was able to rebound at its pretreatment level for viruses harboring one mutation suggest that the genetic barrier of this compound seems to be relatively thin and that the selection of only one mutation is able to lead to a virological fealure to raltegravir. This fact implicates that this compound should be used only with other active drugs that have to be combined to it in the regimen. Finally, most of these mutations led to strongly crippled enzymes. The mechanism by which the virus can support efficient replication while encoding a catalytically inactive integrase remains to be elucidated. Several viral or cellular factors can be evolved in such phenomenon".
ABSTRACT
Raltegravir (MK-0518) is a potent integrase inhibitor that is highly active in patients harboring resistance viruses to other antiretroviral classes. It has been recently shown that raltegravir can select mutations in integrase gene in patients failing to raltegravir regimen.
Four heavily pretreated patients who received a salvage therapy including raltegravir and who subsequently developed virological failure under raltegravir therapy were studied. To describe the genetic changes in the integrase gene, the complete nucleotide sequence of the 864 nucleotides integrase coding region was amplified and determined at different time points during the follow-up of each patient. The 288 corresponding amino acid were deduced, analysed and compared to the point at the beginning of the treatment. The PCR products were cloned into bacterial expression vectors and the 8 proteins produced and purified were assayed in vitro.
Four different profiles of mutations were identified: G140S + Q148H that seemed to appear at the same time; N155H; E92Q and E157Q mutations. In vitro, the four wild-type recombinant integrases were sensitive to raltegravir showing a strand transfer activity specifically inhibited by raltegravir and an unaffected 3' processing activity. All identified mutations altered the functions of integrase protein, E92Q was moderately affected for both 3'processing and strand transfer activities, N155H remained fully active for 3'processing but was markedly impaired for strand transfer, G140S + Q148H was strongly impaired for both activities, retaining only little residual activity and E157Q was almost completely inactive for both 3'processing and strand transfer.
E92Q, G140S + Q148H and N155H integrase mutations, located near the catalytic site of the enzyme, have been identified in patients failing to raltegravir and led to an in vitro resistance. Finally these mutations led to strongly crippled enzymes when tested in vitro in the absence of raltegravir affecting the strand transfer activity in all cases and the 3' processing in some cases.
METHODS, MATERIALS
Four HIV-1 infected patients with virological failure (HIV-1 plasma viral load (VL) > 1000 copies/mL) under MK-0518 containing regimen were analyzed
The integrase gene were amplified and sequenced
Ten microliters of RNA were used for reverse transcription (RT)-PCR (Titan One-Tube RT-PCR kit; Roche Applied Science) following the manufacturer's instructions and using 0,4 µM (final concentration) of each primers IN12 and IN13
The second-round PCR using the AmpliTaq DNA polymerase (Applied Biosystems) was performed following the manufacturer's instructions and using 1 µM (final concentration) each primers IN1 and BH4
The integrase gene was sequenced using a cycle sequencing reaction with the Big Dye terminator kit (Applied Biosystems). A set of 4 primers were used for a complete double-strain coverage of the integrase gene
Production and purification of integrase
The PCR products corresponding to the whole IN sequences were digested with NdeI and BamHI and finally cloned into the pET-15b bacterial expression vector
His-tagged INs were expressed in Escherichia coli BL21 (DE3) and purified under native conditions
IN activity assay
For activity assays, 21-mer ODN U5 (5'-GTGTGGAAAATCTCTAGCAGT-3') was radiolabelled with T4 polynucleotide kinase (Biolabs) and g[32-P]ATP (3000 Ci/mmol) (Amersham), and purified on a Sephadex G-10 column (GE Healthcare). Double-stranded ODN substrate was obtained by mixing equimolar amount of labeled U5B with its complementary strands (5'-ACTGCTAGAGATTTTCCACAC-3') in the presence of 100 mM NaCl. Reactions were carried out for 1 hr at 37°C, in a buffer containing 10 mM HEPES (pH7.2), 1 mM DTT, 7.5 mM MgCl2 in the presence of 12.5 nM dsDNA substrate and recombinant IN at various concentrations. Products were separated by electrophoresis in denaturing 18% acrylamide/urea gels. Gels were analysed with STORM 840TM phosphoimager (GE Healthcare Life sciences, Piscataway, USA) and quantified with ImageQuantTM 4.1 software. IC50 were derived from Inhibition curves fitted to experimental data with Prism 4.0 software (Graphpad software Inc, Sand Diego, USA)
RESULTS
Patients and viral load
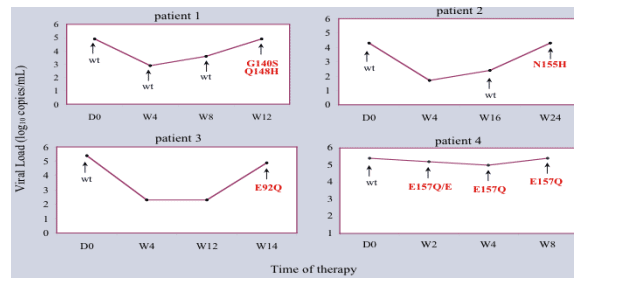
The HIV-1 viral load decreased from baseline to become indetectable (CV<1.69 log10 copies/mL) in 4 weeks, in patients 2 and 3. In the patient 1, the viral load decreased as well but remained detectable (CV=2.86 log10 copie/mL) while the patient 4 showed only a little decrease of the viral load (CV=5.0 log10 copie/mL).
These 4 patients were retrospectively studied when they escaped to the treatment and returned to their initial viral load in a variable time from 8 to 24 weeks
Expression and sensitivity of the wild-type integrases for the 3' processing and the strand transfert activities to MK-0518
As expected, the four wild-type recombinant integrases were very sensitive to MK-0518. IC50s which were derived from in vitro dose-response assays were comparable (7-10nM)
Strand transfer activity was specifically inhibited whereas 3'processing was unaffected for inhibitor concentrations up to 100 fold that of the IC50, thus confirming the mechanism of action postulated for strand transfer inhibitors
Comparison of wild-type and mutant integrases sensitivity to MK-0518
The mutant enzymes were first tested for their catalytic activities. All mutations diversely altered the functions of integrase in vitro. The profile 3 (E92Q) was moderately affected for both 3'processing and strand transfer activities. The profile 2 (N155H) remained fully active for 3'processing but was markedly impaired for strand transfer. The profile 1 (G140S + Q148H) was strongly impaired for both activities, retaining only little residual activity. Finally, the profile 4 (E157Q) was almost completely inactive for both 3'processing and strand transfer. The mutated proteins 1,2 and 3 retained enough activity for testing their sensitivity to MK0518
The three proteins showed a resistance compared to the parental sequences. The profiles 1 and 3 displayed a comparable 7 to 8 fold resistance index while profile 2 was more than 14 fold less sensitive to MK0518. The lack of in vitro activity of profile 4 did not permit the inhibitor to be tested
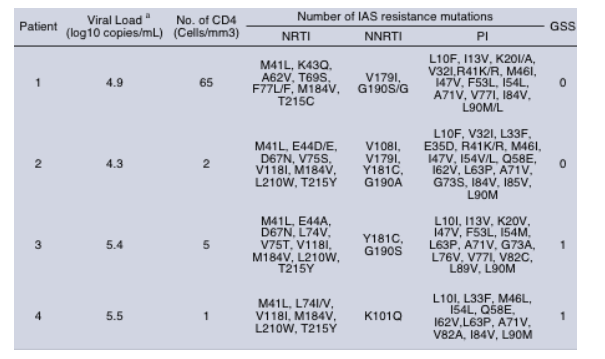
Amino acid substitutions observed from the 4 studied patients during the MK-0518 therapy
This analysis showed a low divergence all along the protein for patients 1, 2, 3 and 4 that represents 4%, 3.8%, 2.7% and 3.8% of variability, respectively
Four different patterns of mutations were identified. The patient 1 showed 2 mutations that seemed to appear in the same time, the G140S and Q148H (profile 1) whereas patients 2,3 and 4 showed only one mutation, N155H (profile 2), E92Q (profile 3) and E157Q (profile 4), respectively
|
| |
|
|
|
|
|