| |
Sustained virologic control in SIV+ macaques after antiretroviral
and α4β7 antibody therapy - possible functional cure
|
| |
| |
Download the PDF here
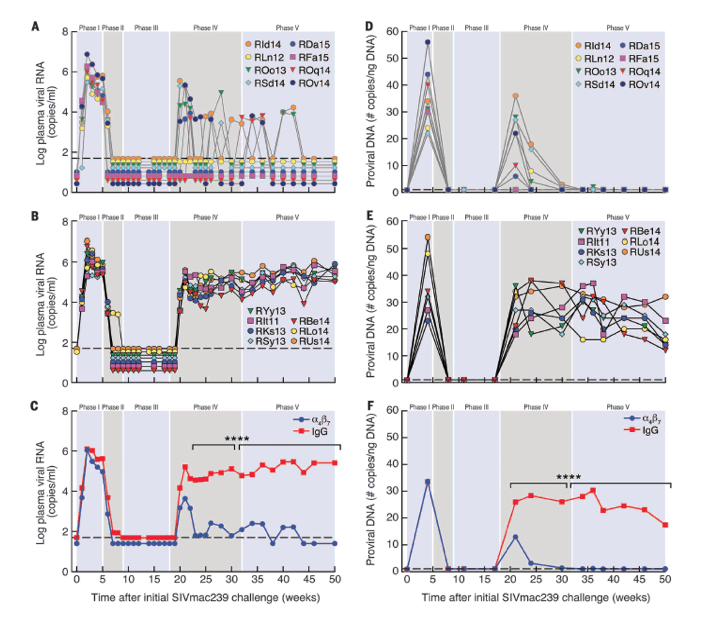
"Immediately after cessation of ART, all monkeys showed increases in proviral DNA. High levels (20 to 40 copies/ng DNA) persisted in the seven IgG-treated animals until week 50 (Fig. 1E). In contrast, in all eight α4β7 mAb-treated macaques, proviral DNA decreased to levels at or below the level of detection by week 30 (Fig. 1D). The difference in the geometric means of the two groups (Fig. 1F) during phases IV and V was significant (P < 0.0001). Undetectable proviral DNA loads in all eight α4β7 mAb-treated macaques persisted well after the final infusion of α4β7 mAb (week 32). Thus, combining α4β7 mAb therapy with short-term ART promoted persistent systemic and mucosal virologic control following discontinuation of all therapy."
"But Picker allows that no other group has yet published similar results. And perhaps most important, unlike the CD4 monoclonal antibody, the one against α4β7 has a human version, vedolizumab, already approved for clinical use. Indeed, NIAID began studies of it in HIV-infected people 3 weeks ago. The trial, which hopes to enroll 20 people, is mainly a safety assessment, but participants will go off ARVs, and then the researchers will closely monitor them to see whether their HIV levels rise or remain suppressed. "We're going to find out very soon whether this is all a bunch of nonsense or actually works," Fauci says."
Link to NIH Study:
https://clinicaltrials.gov/ct2/show/NCT02788175?term=vedolizumab+hiv&rank=1
"Because ART provides only partial protection to GITs (5, 14), we considered the possibility that adding α4β7 mAb might improve this protection. To this end, we conducted a study in genetically characterized (table S1, A to D) SIV-infected RMs that combined a 90-day course of ART, beginning 5 weeks post-infection with a series of eight infusions of α4β7 mAb."
Science Oct 13 2016 - Siddappa N. Byrareddy,1* James Arthos,2* Claudia Cicala,2* Francois Villinger,1,3‡ Kristina T. Ortiz,1 Dawn Little,1 Neil Sidell,4 Maureen A. Kane,5 Jianshi Yu,5 Jace W. Jones,5 Philip J. Santangelo,6 Chiara Zurla,6 Lyle R. McKinnon,7§ Kelly B. Arnold,8 Caroline E. Woody,8 Lutz Walter,9 Christian Roos,9 Angela Noll,9 Donald Van Ryk,2 Katija Jelicic,2 Raffaello Cimbro,10 Sanjeev Gumber,3 Michelle D. Reid,1 Volkan Adsay,1 Praveen K. Amancha,3 Ann E. Mayne,1 Tristram G. Parslow,1 Anthony S. Fauci,2 Aftab A. Ansari1||
1Department of Pathology and Laboratory Medicine, Emory University School of Medicine, Atlanta, GA 30322, USA.
2Laboratory of Immunoregulation, NIAID, NIH, Bethesda, MD 20892, USA.
3Division of Pathology, The Yerkes National Primate Center of Emory University, Atlanta, GA 30329, USA.
4Department of Obstetrics and Gynecology, Emory University School of Medicine, Atlanta, GA 30322, USA.
5Department of Pharmaceutical Sciences, School of Pharmacy, University of Maryland, Baltimore, MD 21201, USA.
6Wallace H. Coulter Department of Biomedical Engineering, Georgia Institute of Technology and Emory University, Atlanta, GA 30680, USA.
7Centre for the AIDS Program of Research in South Africa (CAPRISA), Durban, South Africa.
8Department of Biomedical Engineering, University of Michigan, Ann Arbor, MI 48109, USA.
9Primate Genetics Laboratory, German Primate Center, Leibniz Institute for Primate Research, Gottingen, Germany.
10Division of Rheumatology, Johns Hopkins School of Medicine, Baltimore, MD 21201, USA.
Abstract
Antiretroviral drug therapy (ART) effectively suppresses replication of both the immunodeficiency viruses, human (HIV) and simian (SIV); however, virus rebounds soon after ART is withdrawn. SIV-infected monkeys were treated with a 90-day course of ART initiated at 5 weeks post infection followed at 9 weeks post infection by infusions of a primatized monoclonal antibody against the α4β7 integrin administered every 3 weeks until week 32. These animals subsequently maintained low to undetectable viral loads and normal CD4+ T cell counts in plasma and gastrointestinal tissues for more than 9 months, even after all treatment was withdrawn. This combination therapy allows macaques to effectively control viremia and reconstitute their immune systems without a need for further therapy.
Profound and durable suppression of HIV by antiretroviral therapy (ART) represents a major accomplishment in HIV-AIDS research. However, HIV persists in patients despite long-term ART therapy such that, once ART is withdrawn, virus invariably rebounds. Lifetime ART treatment is associated with toxicity (1), residual chronic inflammation, and the accelerated onset of diseases associated with aging (2, 3).
High levels of viral replication in gastrointestinal tissues (GITs) during acute infection lead to severe depletion of local CD4+ T cells (4), damage to the gut epithelium, and the rapid formation of persistent viral reservoirs. Generalized immune dysfunction and chronic immune activation follow. Even when administered days after infection, ART fails to fully reverse these insults (5). We reasoned that preventing HIV-susceptible cells from accessing GITs might reduce damage to the gut and the mucosal immune system in a way that would allow immune mechanisms to effectively control infection.
A principal pathway that CD4+ T cells use to traffic into GITs involves an interaction between integrin α4β7, expressed on CD4+ T cells, with mucosal vascular addressin cell adhesion molecule 1 (MAdCAM-1), expressed primarily on high endothelial venules within GITs (6, 7). CD4+ T cells that express high levels of the α4β7 integrin (α4β7hi) are preferential targets of HIV and simian immunodeficiency virus (SIV) during acute infection (8-12). In order to disrupt trafficking of α4β7hi CD4+ T cells into GITs, we developed a recombinant rhesus monoclonal antibody against the heterodimeric form of α4β7 (α4β7 mAb) that blocks α4β7 binding to MAdCAM (13-15).
Administration of α4β7 mAb before and during repeated low-dose intravaginal SIV challenge of rhesus macaques (RMs) leads to significant protection from transmission (13). In treated animals that became infected, GIT CD4+ T cells were preserved and GIT proviral DNA was reduced, and thus, virus-mediated damage to GITs was minimized.
Because ART provides only partial protection to GITs (5, 14), we considered the possibility that adding α4β7 mAb might improve this protection. To this end, we conducted a study in genetically characterized (table S1, A to D) SIV-infected RMs that combined a 90-day course of ART, beginning 5 weeks post-infection with a series of eight infusions of α4β7 mAb. This treatment strategy included five phases as outlined in fig. S1. In phase 1 (weeks 1 to 5), 18 RMs were infected intravenously with a 200 median tissue culture infectious dose (TCID50) of SIVmac239. After 5 weeks, all 18 animals began a 90-day daily regimen of ART (phase II). During phase III (weeks 9 to 18), 11 animals received α4β7 mAb once every 3 weeks (eight infusions total); 7 animals received nonspecific rhesus immunoglobulin G (IgG). During phase IV (weeks 18 to 32), ART was withdrawn, and α4β7 mAb-IgG treatment was continued. In phase V (weeks 32 to 50), all treatment was terminated. Three out of 11 α4β7 mAb-treated animals developed antibodies against the α4β7 mAb (fig. S2) and were excluded from further analysis.
Discussion
Combining ART with α4β7 mAb promoted prolonged virologic control (Fig. 1 and fig. S3) and the restoration of CD4+ T cells. Control persisted long after α4β7 mAb treatment was terminated. It was not associated with neutralizing antibody or classical cell-mediated immune responses (figs. S20 to S24) but instead with reduced damage to GITs. The precise mechanism(s) by which ART + α4β7 mAb therapy promoted virologic control remains to be defined. However, we identified a series of correlates that individually or in combination may have contributed to that control. These include the recovery of TH17 and TH22 subsets of CD4+ T cells, significant increases in cytokine-synthesizing NK cells and NKp44+ ILCs, skewing of the antibody response toward the gp120 V2 domain, distinguishing plasma biomarkers in phase III followed by signatures associated with reduced gut damage and inflammation in phases IV and V, and the recovery of RA levels. RA is a key regulator of gut immune responses (31, 32) but it also inhibits fibrosis (33).
Vedolizumab, the humanized analog of α4β7 mAb is believed to reduce trafficking of α4β7-expressing CD4+ T cells to GITs. This is the basis for its use in the treatment of inflammatory bowel disease (IBD) (34). Yet, paradoxically, in this study α4β7 mAb promoted the repopulation of GITs with CD4+ T cells. Future studies are needed to address the phenotype and functionality of these cells. Such information may help us better understand both the virologic control we are observing and the mechanism of action of drugs like vedolizumab. Vedolizumab belongs to a class of therapeutic agents that are currently in various stages of development for the treatment of IBD (34, 35). It may be possible to use these drugs as adjunctive agents in the treatment of HIV infection.
ART + α4β7 mAb controls plasma and gut viral loads
All 15 animals showed similar peaks in viremia around weeks 2 to 3 (∼2.9 x 106 copies/ml), and they all fully suppressed viremia by 3 weeks after ART initiation. The two groups developed divergent viral load (VL) patterns after ART was withdrawn (phase IV). In all seven IgG-treated animals, viremia rebounded to high levels (∼106 copies/ml) within 2 weeks and maintained those levels out to week 50 (Fig. 1B). In contrast, two out of eight α4β7 mAb-treated animals never rebounded, and the remaining six out of eight rebounded but then regained control of viremia within 4 weeks (Fig. 1A). Virologic control was robust in all eight α4β7 mAb-treated animals, with either complete control or transient low-level blips of viremia. The difference in viremia between the two groups after discontinuation of ART was significant (P < 0.0001) (Fig. 1C). Virologic control in all eight α4β7 mAb-treated animals persisted to week 81 (fig. S3), although the last infusion of α4β7 mAb (half-life of ∼11.4 days) was administered at week 32 (13).
Both treatment groups showed similar levels of proviral DNA in GITs during phases I and II. Immediately after cessation of ART, all monkeys showed increases in proviral DNA. High levels (20 to 40 copies/ng DNA) persisted in the seven IgG-treated animals until week 50 (Fig. 1E). In contrast, in all eight α4β7 mAb-treated macaques, proviral DNA decreased to levels at or below the level of detection by week 30 (Fig. 1D). The difference in the geometric means of the two groups (Fig. 1F) during phases IV and V was significant (P < 0.0001). Undetectable proviral DNA loads in all eight α4β7 mAb-treated macaques persisted well after the final infusion of α4β7 mAb (week 32). Thus, combining α4β7 mAb therapy with short-term ART promoted persistent systemic and mucosal virologic control following discontinuation of all therapy.
Rebound of CD4+ T cell subsets
Blood and GIT mononuclear cells isolated from each phase of the study were analyzed by flow cytometry (table S2, A to C). During phase I, the absolute numbers of blood total CD4+ T cells and subsets showed a sharp decline (Fig. 2, A to D). After the first administration of α4β7 mAb (phase III), CD4 values diverged. The α4β7 mAb-treated animals, but not controls, showed marked increases in total CD4+ T cells and, notably, in the effector memory (EM) CD4+ T cell subset (>fourfold increase, P < 0.0001) (Fig. 2D). These increases were sustained after discontinuation of the α4β7 mAb treatment (phase V). By week 50, total CD4+ T cell numbers approached preinfection levels.
Acute HIV-1 and SIV infections are characterized by a rapid depletion of CD4+ T cells in GITs (4, 15-18). Therefore, we evaluated the fate of GIT CD4+ T cells (Fig. 2, E to H), with values expressed as the percent of CD4+ T cells within the gated population of CD45+ cells. We observed a sharp decline in total CD4+ T cells during the acute phase in both groups with CD45+/CD4+ T cells reaching their nadir by the end of phase I (Fig. 2E). In phase III, the CD4+ T cell profile of the two treatment groups diverged. The relative frequency of CD45+/CD4+ cells in the α4β7 mAb-treated animals gradually increased through phase V (P < 0.0001). CD4+ T cell subsets showed an apparent recovery of both T central memory (CM) and T EM cells in the α4β7 mAb-treated animals, with the EM recovering at a faster rate (Fig. 2, G and H). The relative proportion of naïve CD4+ T cells remained constant through phase V (Fig. 2F). The quality of this recovery was well reflected by increases in the frequencies of T helper 17 (TH17) and T helper 22 (TH22) subsets of CD4+ T cells in both GITs and blood (fig. S4) (19). Consistent with these results, immunohistological analysis of GIT sections after week 50 revealed abundant CD4+ T cells in α4β7 mAb-treated animals but not in controls (fig. S5).
To better understand the repopulation of gut tissues with CD4+ T cells, we used a newly developed antibody-targeted positron emission tomography immuno-PET) combined with the computed tomography (CT) imaging technique (20). Around week 50 (phase V), four macaques from each treatment group were imaged with a 64Cu-labeled F(ab')2 antibody against CD4. Although we had originally hypothesized that α4β7 mAb would inhibit CD4+ T cell trafficking to GITs, we instead observed repopulation of CD4+ cells in a wide variety of immune tissues, including GITs (Fig. 3). This result suggests that the protective effect of α4β7 mAb in minimizing GIT viral load early in infection (Fig. 1, D to F) facilitated the repopulation of CD4+ cells throughout the systemic and mucosal immune systems. It is unclear whether the reconstitution of these immune sites resulted from the control of viremia, whether these immune competent components contributed to virologic control, or both.
Phenotypic analysis of NK cells and other cell lineages
Total natural killer (NK) cells in blood remained similar in both groups during phases I to IV (Fig. 4) and decreased during phase V but only in the IgG-treated group (Fig. 4A). We observed a decrease in the cytolytic subset (NKG2a+/CD8+/CD16+/CD56-) during phase III and IV in the α4β7 mAb-treated group, followed by an increase during phase V (Fig. 4B). Cytokine-synthesizing NK cells (NKG2A+/CD8+/CD16-/CD56+) increased during phase IV in α4β7 mAb-treated animals (Fig. 4C). By week 50, these values approached baseline. A related pattern was observed in GITs. In the α4β7 mAb-treated group, the proportion of the cytokine-synthesizing NK cell subset increased through phases IV and V (Fig. 4H), with corresponding decreases in the proportion of other NK cell subsets. These changes are noteworthy in two ways. First, among all NK cell subsets analyzed, α4β7 expression is highest (∼40%) on cytokine-synthesizing NK cells. Second, these increases coincided with the appearance of virologic control in phase IV. Frequencies of NKp44+ innate lymphoid cells (ILCs) decreased in both treatment groups as early as phase I (fig. S6), which is consistent with the loss of ILCs in acute HIV infection (21). However, we observed a sustained increase in the frequency of ILCs in gut biopsies in ART + α4β7mAb-treated animals beginning in phase III but not in controls. Note that vaccine-induced ILCs have been correlated with protection from SIV infection (22). Other cell lineages (CD8+ T cells, B cells, and plasmacytoid and myeloid dendritic cells) were analyzed (figs. S7 and S8), as were activation markers on CD4+,CD8+ T and NK cells (figs. S9 to S11). Although differences were noted, further studies will be required to inform the impact of those differences.
Identification of signature plasma cytokines
Results presented above suggest that virologic control was immune mediated. To this end, we analyzed a panel of 20 immune or inflammatory markers (table S3) in plasma samples. To identify signatures for each of the five phases of the study, we used a partial least square discriminant analysis (PLSDA). PLSDA models were created for the five study phases by using measurements from each sample and time point, and variable importance projection scores were used to omit markers that did not contribute to group differentiation. Cross-validation was used to assess model performance. In phases I and II, we found no distinction between treatment groups, with a high cross-validation error between models (0.28 and 0.21, respectively) (fig. S12, A and B). In phases III to V, distinct signatures differentiated the two treatment groups, with low cross-validation errors in each phase (0.09, 0.02, and 0, respectively) (Fig. 5A). The differentiating signature varied in each of these phases. One common feature of all three was a comparative increase in retinoic acid (RA) in the α4β7 mAb-treatment group (Fig. 5B). The signature in phase III, when both groups were aviremic, included increases in interleukin-21 (IL-21), granulocyte-macrophage colony-stimulating factor, soluble CD14 (sCD14), interferon-α (IFN-α), as well as RA; however, IFN-Υ and transforming growth factor-β (TGF-β) were reduced. In phases IV and V, α4β7 mAb treatment was associated with increased IL-10 and RA, with comparative decreases of proinflammatory markers, including IL-1β, IFN-Υ-induced protein (IP-10), complement-reactive protein (CRP), the coagulation biomarker D-dimer, and two markers, sCD163 and intestinal fatty acid-binding protein (I-FABP), associated with gut permeability (Fig. 5A and fig. S13).
ART + α4β7 mAb restores plasma retinoic acid levels
RA induces the expression of α4β7 (23, 24) and also plays an essential role in gut homeostasis (23, 25-27). Baseline RA levels decreased during acute infection in both groups (Fig. 5B). A similar response was observed in two uninfected monkeys afflicted with chronic diarrhea (fig. S14), suggesting that such declines are linked to gut inflammation. Of note, measurements taken on weeks 15, 17, and 19 (phase III) differentiated the two groups, such that RA in the α4β7 treatment group recovered to near-baseline levels. To understand whether these changes reflected the administration of α4β7 mAb, independent of SIV infection, we treated two uninfected animals with α4β7 mAb and observed no effect on serum RA (fig. S15). Indeed, we also observed a similar response in I-FABP levels in phase III (fig. S13). Finally, we carried out an exploratory analysis to identify additional biomarkers in phase III that might correlate with the degree of gut proviral DNA rebound in phase IV. Both positive and negative predictors of the magnitude of viral rebound in gut tissues were identified (fig. S16).
Induction of gp120 V2-specific antibody responses
Plasma and peripheral blood mononuclear cell (PBMC) samples were also screened for SIV-specific antibody responses and antibody-dependent cell-mediated cytotoxicity (ADCC). We found neither neutralizing antibodies nor differences in ADCC titers (fig. S17). In human and macaque vaccine trials, nonneutralizing antibodies directed against the second variable loop (V2) in Env glycoprotein 120 (gp120) correlated with reduced risk of acquisition (28, 29). We therefore characterized the specificity of the antibody responses by performing Pepscan enzyme-linked immunosorbent assay (ELISA) of sera from five animals in each group against linear peptides spanning the entire gp120 subunit (fig. S18). Serum samples from weeks 30 to 36 showed that five out of five α4β7 mAb-treated macaques reacted, to varying levels, with two overlapping V2 peptides (peptides 26 and 27). None of the IgG-treated controls reacted to both of these peptides (two animals reacted to one peptide). We found no other consistent difference in epitope-specific reactivity between the two groups. We then evaluated antibody responses using a more sensitive surface plasmon resonance (SPR) assay. Sera from all 15 animals through the five treatment phases were reacted with both SIV gp120 and an SIV cyclic V2 peptide. Sera from both treatment groups showed similar reactivity to gp120 (mean values) (fig. S19). However, the two treatment groups differed in their reactivity to V2. Whereas eight out of eight α4β7 mAb-treated animals showed persistent reactivity to V2, only three out of seven IgG-treated animals reacted to V2 (Fig. 6A). We then fine-mapped this reactivity by competing serum reactivity from all eight animals with overlapping 15mer peptides spanning V2 (Fig. 6B). A peptide corresponding to the sequence KFNMTGLKRDKKKEY (see Fig. 6 legend) reduced serum reactivity by ∼80%. Alignment of this sequence with an HIV Thai clade A/E V2 sequence indicates that it recognizes the same region identified in a sieving analysis for immune correlates of reduced risk in the RV144 vaccine trial (Fig. 6C) (30). These data indicate that α4β7 mAb treatment promotes V2 antibody responses by an undefined mechanism.
|
|
| |
| |
|
|
|