| |
Prevalence of CXCR4 Tropism among Antiretroviral-Treated HIV-1-Infected Patients with Detectable Viremia
|
| |
| |
The Journal of Infectious Diseases Oct 1, 2006;194:926-930
Peter W. Hunt,1 P. Richard Harrigan,10,11 Wei Huang,9 Michael Bates,9 David W. Williamson,3,8 Joseph M. McCune,4 Richard W. Price,2 Serena S. Spudich,2 Harry Lampiris,5 Rebecca Hoh,1 Teri Leigler,1 Jeffrey N. Martin,1,6,7 and Steven G. Deeks1
1Positive Health Program and 2Department of Neurology, San Francisco General Hospital, 3Graduate Program in Biological and Medical Informatics, 4Division of Experimental Medicine, Department of Internal Medicine, 5San Francisco Veterans Affairs Medical Center, 6Center for AIDS Prevention Studies, and 7Department of Epidemiology and Biostatistics, University of California, San Francisco, 8Gladstone Institute of Virology and Immunology, San Francisco, and 9Monogram Biosciences, Inc., South San Francisco, California; 10British Columbia Centre for Excellence in HIV/AIDS, Providence Health Care, and 11Faculty of Medicine, University of British Columbia, Vancouver, Canada
"....Although much is known about the prevalence of CXCR4 tropism among patients with untreated HIV-1 infection, comparatively little is known about the role that CXCR4 tropism plays in treated patients with drug-resistant viremia. In the present study, we demonstrate that treated participants with detectable viremia, regardless of antiretroviral regimen, have 4-fold greater odds of dual/mixed/X4 tropism than treatment-naive participants, independent of CD4+ T cell count and CCR5 delta32 genotype. Although most of this enrichment for dual/mixed/X4-topic viruses appears to be explained by lower pretreatment nadir CD4+ T cell counts, X4-tropic variants apparently persist despite treatment-mediated restoration of peripheral CD4+ T cell counts...."
Abstract
Although CXCR4-tropic viruses are relatively uncommon among untreated human immunodeficiency virus (HIV)-infected individuals except during advanced immunodeficiency, the prevalence of CXCR4-tropic viruses among treated patients with detectable viremia is unknown. To address this issue, viral coreceptor usage was measured with a single-cycle recombinant-virus phenotypic entry assay in treatment-naive and treated HIV-infected participants with detectable viremia sampled from 2 clinic-based cohorts. Of 182 treated participants, 75 (41%) harbored dual/mixed or X4-tropic viruses, compared with 178 (18%) of the 976 treatment-naive participants (P < .001). This difference remained significant after adjustment for CD4+ T cell count and CCR5 delta32 genotype. Enrichment for dual/mixed/X4-tropic viruses among treated participants was largely but incompletely explained by lower pretreatment nadir CD4 + T cell counts. CCR5 inhibitors may thus be best strategically used before salvage therapy and before significant CD4 + T cell depletion.
With CCR5 inhibitors, a new class of antiretroviral medications targeting HIV-1 entry, in phase 3 clinical trials, there is renewed interest in the prevalence of viruses capable of using the alternative CXCR4 coreceptor (X4) for entry. Among untreated patients, R5-tropic viruses predominate during most stages of HIV infection [1], accounting for 82% of isolates from treatment-naive patients initiating antiretroviral therapy in a recent population-based study [2]. However, the prevalence of viruses that can use the CXCR4 coreceptor for entry approaches almost 50% among untreated individuals with advanced immunodeficiency [1, 2].
Comparatively little is known about the prevalence of X4-tropic viruses in antiretroviral-treated patients with detectable viremia. Because CCR5 inhibitors are likely to be initially used in treated patients with resistance to currently available antiretroviral medications, it is important to assess the prevalence of CXCR4 tropism in this population. There are several reasons to believe that antiretroviral therapy might alter the prevalence of X4-tropic viruses. First, partially suppressive therapy may lead to an increase in HIV-specific T cell responses [3]. Because X4-tropic variants may be more susceptible to cytotoxic T cell responses than R5-tropic viruses [4], increases in HIV-specific T cell responses during partial treatment-mediated viral suppression might select against X4-tropic viruses. Second, antiretroviral therapy reduces CCR5 expression on T cells, presumably as a consequence of reductions in T cell activation [5, 6], potentially selecting for X4-tropic viruses [2]. Last, certain antiretroviral drugs may preferentially select for one virus population, either because of enhanced activity against X4 viruses (as has been suggested for enfuvirtide [7]) or because of suboptimal drug metabolism in the cellular reservoirs for X4 viruses (as has been suggested for zidovudine [8]). To assess the potential impact that partial treatment-mediated viral suppression has on the prevalence of CXCR4 tropism, we compared the prevalence of X4-tropic viruses between treatment-naive participants and treated participants with detectable plasma HIV RNA levels.
Participants and methods. Antiretroviral-treated participants were sampled from the Study of the Consequences of the Protease Inhibitor Era (SCOPE), an ongoing clinic-based cohort of >600 chronically HIV-infected patients in San Francisco. Participants are seen every 4 months, at which time an extensive evaluation is performed and biologic specimens are obtained. Additional treated participants were sampled from a previously reported cross-sectional study comparing tropism of plasma and cerebrospinal fluid viral isolates [9]. Treated participants were eligible for the current analysis if they were receiving a stable antiretroviral regimen for >4 months and had detectable plasma HIV RNA levels (>50 copies/mL). Participants with exposure to coreceptor antagonists were excluded. Antiretroviral-naive participants were sampled, before starting antiretroviral therapy, from the Highly Active Antiretroviral Therapy Observational Medical Evaluation and Research (HOMER) cohort, a population-based cohort of HIV-1-infected patients within the British Columbia Centre for Excellence in HIV/AIDS network. Coreceptor tropism was measured, in all of these antiretroviral-naive participants with detectable plasma HIV RNA levels, as reported elsewhere [2]. Informed consent was obtained from all participants, and ethics approval was obtained from the ethics boards of each institution.
Plasma HIV RNA levels were determined by the branched DNA (bDNA) amplification technique (Quantiplex HIV RNA, version 3.0; Chiron) for SCOPE participants and by polymerase chain reaction (PCR) (Cobas Amplicor HIV-1 Monitor Test, version 1.5; Roche Diagnostics) for HOMER participants. For treated participants, the pretreatment nadir CD4+ T cell count was the lowest self-reported value before the initiation of the current regimen. For treated participants, resistance to protease inhibitors and reverse transcriptase inhibitors was measured within 6 months of the coreceptor tropism measurement, using the TRUGENE HIV-1 Genotyping Kit (Bayer HealthCare Diagnostics); was analyzed with OpenGene software (Visible Genetics); and was interpreted on the basis of 2004 International AIDS Society-USA guidelines [10].
CCR5 delta32 genotype was determined by extracting DNA from whole blood or peripheral-blood mononuclear cells by use of the QIAamp Blood Midi Kit (Qiagen). Extracted DNA was amplified in a single round of PCR using primers flanking the CCR5 32 region (forward, 5-TCAAAAAGAAGGTCTTCATTACACC-3; reverse, 5-AGCCCAGAAGAGAAAATAAACAATC-3). PCR products were visualized by electrophoresis on a 3% agarose gel and classified on the basis of fragment length (241 bp for wild type and 209 bp for the CCR5 delta32 allele).
The PhenoSense HIV-entry assay (Monogram Biosciences) was used to determine HIV-1 coreceptor use [9]. Briefly, participant-derived env DNA (gp160) was amplified by PCR from plasma isolates and ligated into pCXAS expression vectors. A replication-defective retroviral vector containing a luciferase expression cassette inserted within the env gene was used to cotransfect human embryonic kidney cell cultures with the sample plasmid DNA. Recombinant viruses were harvested after 48 h and were assessed for their ability to infect cells expressing CCR5 or CXCR4 by measuring luciferase activity in the presence of coreceptor-specific inhibitors. The PhenoSense assay classifies isolates as R5-, X4-, or dual and/or mixed-tropic virus.
Plasma HIV RNA levels obtained by use of Cobas Amplicor (version 1.5) were converted to bDNA (Quantiplex HIV RNA, version 3.0) equivalents by subtracting 0.3 log10 copies/mL [11]. For analysis, tropism was dichotomized as either R5- or dual/mixed/X4-tropic, given the small number of purely X4-tropic isolates. Among treated participants, the treatment-mediated change in CD4+ T cell count was defined as the difference between the current CD4+ T cell count (at the time of tropism measurement) and the pretreatment nadir CD4+ T cell count. Factors associated with tropism were assessed in unadjusted analyses with Fisher's exact tests and in adjusted multivariable logistic regression or stratified analyses. Both current and pretreatment nadir CD4+ T cell counts, plasma HIV RNA levels, and CCR5 32 genotype were considered as potential confounding factors/mediators. Backward stepwise model selection was used for multivariable logistic regression models, retaining factors that altered the association between treatment status and tropism by at least 10%.
Results. Compared with the 976 treatment-naive participants, the 182 treated participants had lower plasma HIV RNA levels and pretreatment nadir CD4+ T cell counts, but the majority in each group had a current CD4+ T cell count >250 cells/mm3 (table 1). A similar percentage of participants in each group were heterozygous for the CCR5 delta32 mutation. Most of the treated participants were receiving a protease inhibitor-based regimen and had a moderate number of drug-resistance mutations. Only 10% of treated participants were receiving the HIV-entry inhibitor enfuvirtide.
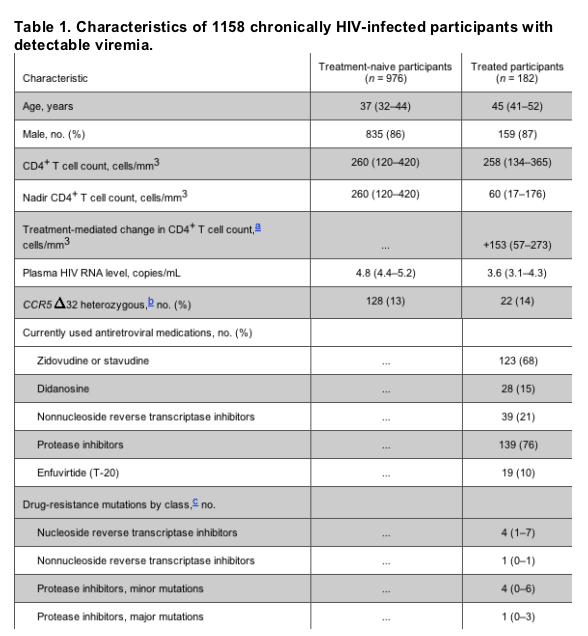
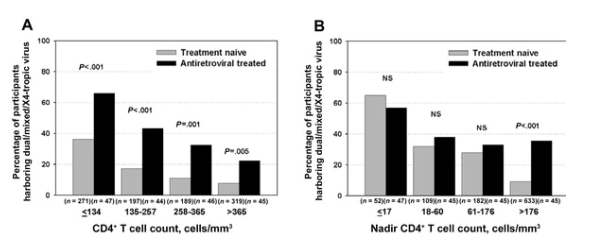
Compared with treatment-naive participants, a higher percentage of treated participants were harboring dual/mixed/X4-tropic viruses (41% vs. 18%; P < .001). Only 1 participant in each group apparently harbored a purely X4-tropic virus population, so these were included with participants harboring dual/mixed-tropic viruses in subsequent analyses. Although the prevalence of dual/mixed/X4 tropism was higher at lower CD4+ T cell counts (P < .001 for trend), the treated participants had a higher prevalence of dual/mixed/X4 tropism at any given CD4+ T cell count (P < .05 within each CD4+ T cell count stratum) (figure 1A).
Figure 1. Prevalence of dual/mixed/X4 tropism by treatment status and either current or pretreatment nadir CD4+ T cell count. The percentage of participants harboring dual/mixed or X4 tropic viruses is plotted by quartiles of current CD4+ T cell count and pretreatment nadir CD4+ T cell count among 976 treatment-naive (gray bars) and 182 treated (black bars) participants with chronic HIV infection. A, Higher prevalence of dual/mixed/X4 tropism at lower current CD4+ T cell counts (P < .001 for trend) and higher prevalence of dual/mixed/X4 tropism for treated participants at any given current CD4+ T cell count (P < .01 within each CD4+ T cell count stratum). B, Association of lower pretreatment nadir CD4+ T cell counts with a higher prevalence of dual/mixed/X4 tropism for both treatment-naive (P < .001 for trend) and treated participants (P = .03 for trend). Among participants with CD4+ T cell count nadirs in the lowest 3 quartiles (176 cells/mm3), there was no evidence for an association between treatment status and dual/mixed/X4 tropism after controlling for pretreatment nadir CD4+ T cell count. However, among participants with pretreatment nadir CD4+ T cell counts >176 cells/mm3, treatment was independently associated with a higher prevalence of dual/mixed/X4 tropism (P < .001). P < .001 for interaction. NS, not significant.
Among 150 participants heterozygous for the CCR5 delta32 mutation, 48 (32%) harbored dual/mixed/X4-tropic virus, compared with only 193 (20%) of 976 participants without this mutation (P = .001). In unadjusted analyses, higher plasma HIV RNA levels were associated with dual/mixed/X4 tropism among treatment-naive participants (P < .001) and treated participants (P = .05). However, among either treatment-naive or treated participants, there was no longer evidence for an independent association between plasma HIV RNA levels and coreceptor tropism after adjusting for CD4+ T cell count (P> .85, for each association). Even after adjusting for current CD4+ T cell count and CCR5 delta32 genotype, antiretroviral-therapy use continued to be associated with 4-fold increased odds of dual/mixed/X4 tropism (95% confidence interval, 2.7-fold to 5.8-fold; P < .001).
Lower pretreatment nadir CD4+ T cell counts were also associated with a higher prevalence of dual/mixed/X4 tropism for both treatment-naive (P < .001 for trend) and treated participants (P = .03 for trend) (figure 1B). After stratifying by pretreatment nadir CD4+ T cell count, there was no longer any evidence for an independent association between antiretroviral therapy and dual/mixed/X4 tropism among those with pretreatment nadir CD4+ T cell counts in the lowest 3 quartiles (<176 cells/mm3) (P > .47 within each stratum). Thus, the enrichment for dual/mixed/X4 tropism among the majority of treated participants in our sample was largely explained by low pretreatment nadir CD4+ T cell counts, even though the nadir had occurred a median of 5 years earlier (interquartile range [IQR], 1-7 years) and CD4+ T cell counts had increased by a median of 148 cells/mm3 (IQR, +79 to +257 cells/mm3) during that time. However, among those with pretreatment nadir CD4+ T cell counts in the highest quartile (>176 cells/mm3), treatment continued to be associated with a higher prevalence of dual/mixed/X4 tropism, independent of the pretreatment nadir CD4+ T cell count (P < .001). This difference remained significant even if the analysis was restricted to those with nadirs <350 cells/mm3 (P < .001).
We next assessed whether the enrichment for dual/mixed/X4-tropic viruses among treated participants might be explained by exposure to specific drugs. Prior studies indicated that thymidine analogs (zidovudine or stavudine) select for syncytium-inducing (and presumably CXCR4-using) viruses in vivo, because these drugs are less likely to be phosphorylated in the cellular reservoirs for these viruses [8]. However, among the 182 antiretroviral-treated participants, we found no evidence for an association between current thymidine-analog use and tropism (P = .63). Enfuvirtide may also interfere with gp120 binding to CXCR4 but not CCR5, potentially selecting for R5-tropic variants [7]. Although underpowered, our study provided no evidence for an association between enfuvirtide use and tropism in our treated participants (P = .63).
Discussion. Although much is known about the prevalence of CXCR4 tropism among patients with untreated HIV-1 infection, comparatively little is known about the role that CXCR4 tropism plays in treated patients with drug-resistant viremia. In the present study, we demonstrate that treated participants with detectable viremia, regardless of antiretroviral regimen, have 4-fold greater odds of dual/mixed/X4 tropism than treatment-naive participants, independent of CD4+ T cell count and CCR5 delta32 genotype. Although most of this enrichment for dual/mixed/X4-topic viruses appears to be explained by lower pretreatment nadir CD4+ T cell counts, X4-tropic variants apparently persist despite treatment-mediated restoration of peripheral CD4+ T cell counts.
Our estimate of an unusually high prevalence of dual/mixed/X4 tropic viruses among treated participants with detectable viremia in the present study is consistent with other recent reports. For example, one-third to one-half of treatment-experienced patients enrolling in clinical trials harbor dual/mixed/X4-tropic viruses [12, 13]. Also, despite previous reports of rare CXCR4 tropism among untreated individuals infected with HIV-1 subtype C, 50% of treated patients infected with drug-resistant HIV-1 subtype C harbored viruses capable of using CXCR4 for entry in one small study [14].
It remains unclear why treated patients with drug-resistant viremia remain enriched for dual/mixed/X4-tropic viruses despite treatment-mediated gains in peripheral CD4+ T cell counts. The association between CCR5 delta32 heterozygosity and dual/mixed/X4 tropism suggests a potential role of CCR5+ target cell availability [2]. Persistent loss of CCR5+ T cell targets during antiretroviral therapy might be explained by poor reconstitution of gut-associated lymphoid tissue, where most CCR5+ T cell targets reside, despite robust peripheral treatment-mediated CD4+ T cell gains [15]. Alternatively, treatment-mediated viral suppression may cause declines in CCR5 expression [5, 6], potentially explaining the enrichment for dual/mixed/X4-tropic viruses observed among participants with high pretreatment nadir CD4+ T cell counts.
This study has some limitations that deserve comment. First, because the tropism measurement used in the present study cannot distinguish between mixtures and dual-tropic viral populations or quantify the relative proportion of X4-tropic viruses within a given dual/mixed sample, it is possible that the high prevalence of dual/mixed-tropic viruses in treated participants is driven by participants with clinically insignificant amounts of X4-tropic virus replication. However, luciferase activity in CXCR4-expressing cells was comparable among dual/mixed-tropic isolates from both groups (data not shown). In addition, because this study was cross-sectional and potentially susceptible to selection bias, we can only speculate as to the timing and causes of tropism switches among treated participants. Longitudinal studies of participants initiating antiretroviral therapy before the onset of virologic failure will be necessary to specifically address these issues.
In summary, we have observed an unexpectedly high prevalence of dual/mixed/X4 tropism among treated participants with detectable viremia. Consequently, treated participants with drug resistance may be less likely than treatment-naive participants to achieve viral suppression while receiving CCR5 inhibitors. If these results are corroborated by other studies, CCR5 inhibitors may be better strategically used before salvage therapy and before advanced immunodeficiency.
|
|
| |
| |
|
|
|