 |
|
|
| |
Inhibitory activity of TMC435350 on HCV NS3/4A
proteases from genotypes 1 to 6
|
| |
| |
Reported by Jules Levin
AASLD Nov 4 2008 San Francisco CA
Tse-I Lin1, Benoit Devogelaere1, Oliver Lenz1, Origene Nyanguile1, Elisabeth van der Helm1, Katrien Vermeiren1, Genevieve Vandercruyssen1
Erna Cleiren1, Jimmy Lindberg2, Michael Edlund2, Pierre Raboisson1, Herman de Kock1, Maxwell Cummings1, Gregory Fanning1, Kenneth Simmen1
1Tibotec BVBA, Mechelen, Belgium; 2Medivir AB, Huddinge, Sweden
AUTHOR CONCLUSION
TMC435 is a potent inhibitor of NS3/4A protease from genotype 1 to 6, with IC50 values below 13 nM for all HCV NS3/4A enzymes tested with the exception of genotype 3a protease (37 nM).
Analysis suggests that residue diversity at D168 is pivotal for reduced activity of TMC435 observed against genotype 3a protease.
TMC435 binds to the protease non- covalently, with a fast association rate and a slow dissociation rate. The dissociative half-life (t1/2) of TMC435 is > 2 min.
In vitro potency, in conjunction with its pharmacokinetic properties in humans, suggests that TMC435 may provide benefits as an antiviral agent for individuals infected with non-genotype-1 HCV.
INTRODUCTION
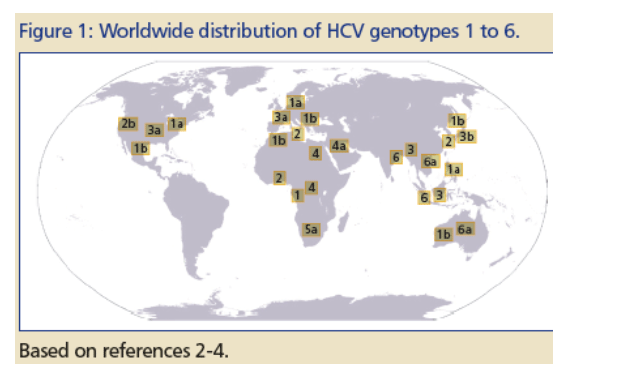
Hepatitis C Virus (HCV; Figure 1):
--120-170 million people infected;·
--displays high degree of genetic variability;
--six genotypes;
--more than 50 subtypes;
--cure rates with current standard of care (pegylated interferon + ribavirin):
~40% for geno-types 1 and 4, about 80% for genotype 2, 3 and 6, and about 60% for genotype 5.(1,2)
TMC435 (formerly known as TMC435350; Figure 2):
--Macrocyclic HCV NS3/4A protease inhibitor.
--EC50 = 8 nM on genotype 1b replicon.
--Limited effect of functional plasma protein binding (about 2-fold shift of EC50 value).
--Currently in Phase 2a of clinical development
Here we present the activity of TMC435 on a panel of genotype 1 to 6 NS3 enzymes obtained from patient isolates using biochemical protease assays and the kinetic binding analysis of genotype 1 isolates with surface plasmon resonance (SPR) based technology.
RESULTS
Inhibitory activity of TMC435 against NS3 proteases of genotype 1 to 6
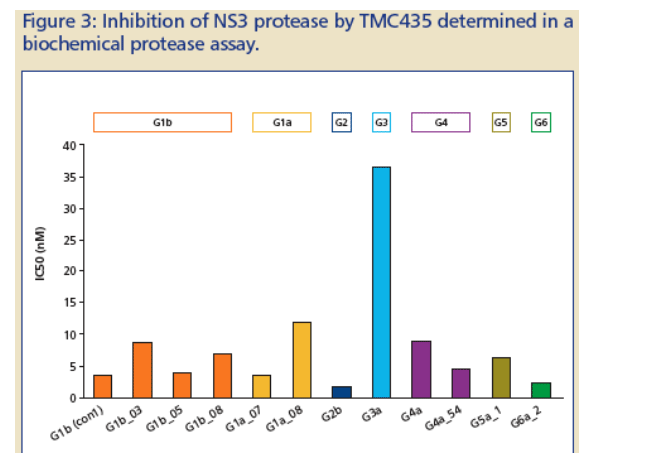
TMC435 activity on NS3 proteases derived from clinical isolates of genotypes 1a, 1b, 2b, 3a, 4a, 5a and 6a (Figure 3, Table 1 ):
-- IC50 values for genotype 1a, 1b, 2b, 4a, 5a, and 6a proteases were less than 5-fold higher than the genotype 1b (con1) values.
-- For genotype 3a protease, a about 10-fold reduction in inhibitory activity, compared to genotype 1, was observed.
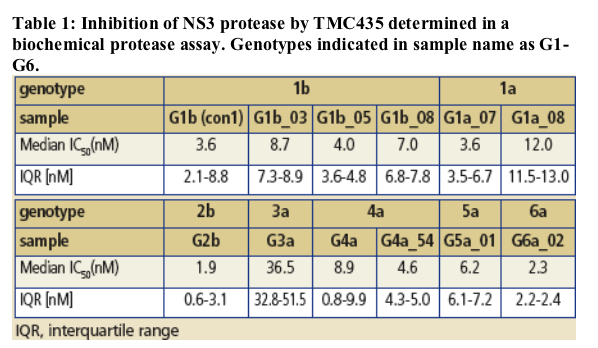
NS3 proteases were cloned from clinical isolates of HCV genotype 1 to 6, expressed and purified from E. coli. The IC50 value for protease inhibition by TMC435 was determined by measuring the fluorescence resonance energy transfer (FRET) induced by the cleavage of the RetS1 peptide substrate when incubated with NS3 protease domain (supplemented with synthetic truncated NS4A peptide).
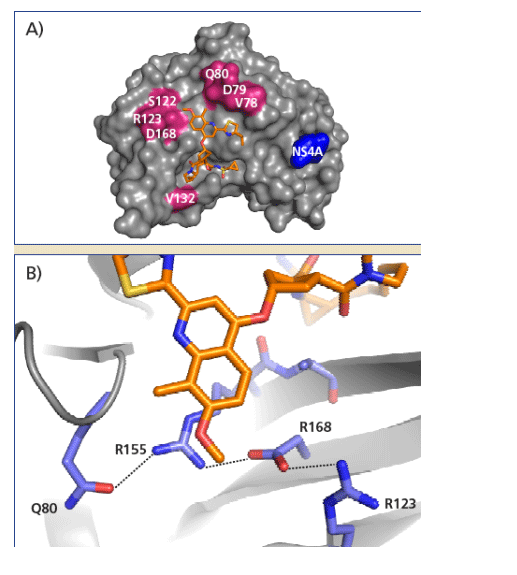
Genotype dependant difference of TMC435 binding pocket residues
Several residue positions in the region of the TMC435 binding site of NS3 show variation between different genotypes (Table 2, Figure 4A ).
The reduced activity against genotype 3 enzymes can be explained by the predominant presence of D168Q variation in this genotype. Variations at position 168 can have a significant impact on the affinity of NS3 protease for TMC435 (Figure 4B).
-- D168 plays a central role in a network of · interactions that stabilizes the shape of the S2 pocket that is induced by TMC435 binding.
-- Mutations of D168 will disturb this interaction · network, through loss of stabilizing hydrogen bonds and/or by introduction of steric clashes.
Depending on the residue at position 168, signifi-cant alteration of the overall shape and chemical nature of the S2 region that binds the methoxy-quinoline-group of TMC435 can be expected.
The D168E variation present in genotype 5a is conservative and hence allows preservation of intramolecular contacts.
Analysis of a single D168E or D168Q mutation in a genotype 1b background using a transient replicon assay confirmed the greater impact of D168Q on susceptibility to TMC435 compared to D168E, suggesting that the presence of D168Q in the genotype 3 protease might be pivotal for the reduced activity of TMC435 observed against the genotype 3 enzyme.
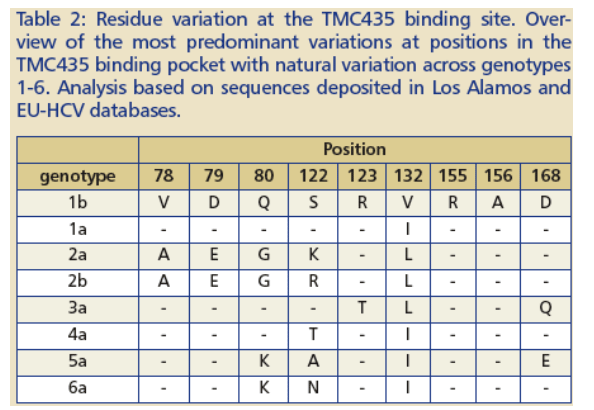
Figure 4: Residue variation at the TMC435 binding site. (A) Crystal structure of TMC435 bound to HCV NS3 protease, showing residue positions in TMC435 binding pocket with natu-ral variation between genotypes 1-6. The observed variable sites are in the vicinity of the S2 or S3 regions of the binding site. (B) Detailed view of the interactions between Q80, R155, D168, R123 and the methoxyquinoline group of TMC435.
Binding of TMC435 to NS3 proteases of genotype 1
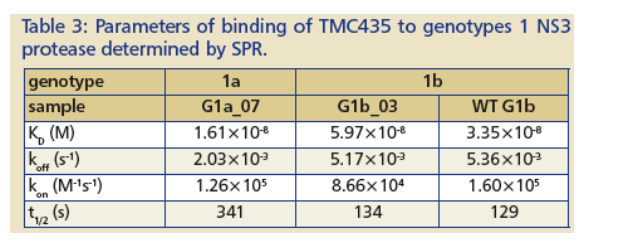
TMC435 binding to NS3 (Figure 5, Table 3) was consistent with:
-- The similar IC· 50 values for genotype 1a and 1b samples (Figure 3).
-- Non-covalent binding to the protease, with a fast association rate (k-on about 10-5th M -1, s-1) and a slow dissociation rate (k-off aout 10 -3, s-1). The dissociative half-life (t1/2) of TMC435 is > 2 min.
-- One-step binding kinetics (data not shown)
Figure 5: Binding of TMC435 to genotypes 1 NS3 protease determined by SPR. (A) Experimental sensorgram showing the interaction between TMC435 (concentration of TMC435 used: 15 to 792 nM) and genotype 1b (con1) NS3. (B) Interaction kinetic map of genotypes 1 NS3 proteases with TMC435, shown as association versus dissociation rate constants (kon and koff), and the combinations of kon and koff that result in the same KD values (diagonal lines).
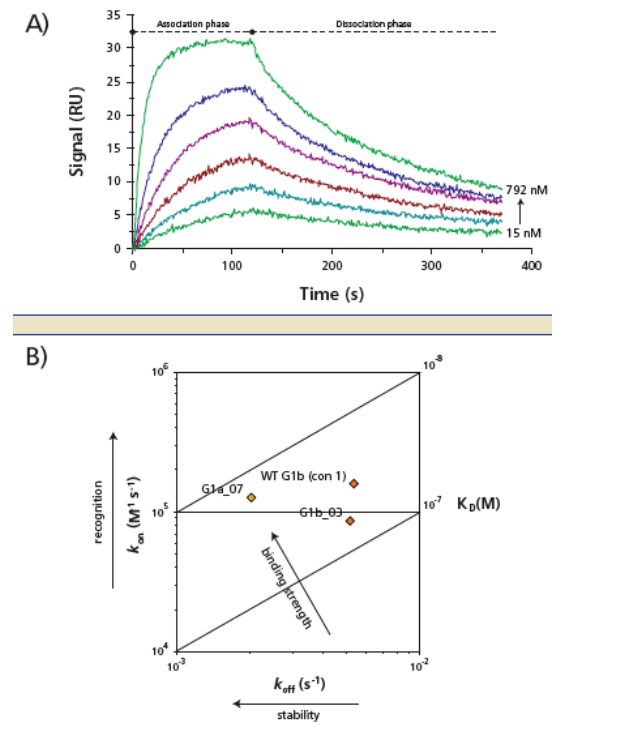
NS3 proteases were cloned from genotype 1 clinical isolates or from strain con1b, expressed and purified from E. coli. To charac-terize the binding of TMC435 to HCV protease, binding kinetics to immobilized recombinant genotype 1 NS3 protease domain was determined using SPR on a Biacore S51 (GE Healthcare).
REFERENCES
1. Strader DB et al. 2004. Hepatology 39: 1147.
2. Nguyen MH and Keeffe EB. 2005. Clin Gastroenterol Hepatol 3: S97.
3. Ramia S and Eid-Fares J. 2006. Int J Infect Dis 10: 272.
4. Simmonds P. 2004. J Gen Virol 85 (Pt 11): 3173-3188.
|
| |
|
|
|
|
|