| |
Keeping Blood Sugar Low May Help Stem Geriatric Memory Loss
|
| |
| |
"Higher levels of blood glucose were related to dysfunction in the dentate gyrus, indicating that blood sugar could be a pathogenicmechanism for cognitive decline."
"elevations in blood glucose as a contributing cause of age-related memory decline, and suggest specific interventions that can preserve cognitive health."
MedPage Today
Published: December 30, 2008
NEW YORK, Dec. 30 -- Lowering blood glucose levels may help lessen the cognitive decline of normal aging, even in diabetes-free patients, researchers here said.
Action Points
* Explain to interested patients that the results of this study suggested that keeping blood glucose levels low through physical activity or other means may slow age-related memory loss.
They sought to see whether lessening blood glucose levels, which have been associated with less activity in the dentate gyrus (P=0.032), could keep cognitive activity from declining, Scott Small, M.D., of Columbia, and colleagues said in the December issue of the Annals of Neurology.
Previous studies have shown that dysfunction in this area of the hippocampus with normal aging contributes to cognitive decline. So Dr. Small and colleagues did MRI scans of 240 apparently dementia-free community-based persons 65 and older (mean age 79.7) and found apparent indications that lower is better.
Higher levels of blood glucose were related to dysfunction in the dentate gyrus, indicating that blood sugar could be a pathogenic mechanism for cognitive decline.
"Our results suggest that improving glucose metabolism is a clinically tractable approach for ameliorating the cognitive slide that occurs in all of us as we age," Dr. Small and colleagues wrote in the Annals of Neurology.
Previous studies have shown that the hippocampus is affected by multiple diseases that are common in late life, such as Alzheimer's disease, stroke, and diabetes.
Because diabetes and infarcts are typically comorbid, the researchers set out to discover how the two conditions influence age-related cognitive decline.
A quarter of the participants who had an MRI had diabetes, and 34.3% had MRI-documented brain infarcts.
Both diabetes (P=0.0004) and stroke (P=0.025) were linked to global hippocampal dysfunction, but the conditions each affected different areas of the hippocampus. Diabetes was linked to dysfunction in the dentate gyrus (P=0.0002).
In addition, higher levels of blood glucose were associated with lower scores on the Selective Reminding Test (P=0.018), a cognitive measure of hippocampal function.
The results were confirmed by imaging aging Rhesus monkeys and a mouse model of hyperglycemia.
Stroke was linked to dysfunction in the CA1 subfield (P=0.036) and the subiculum (P=0.008). The association with the CA1 subfield indicates that transient hypoperfusion could be a pathogenic mechanism for hippocampal dysfunction in patients with vascular strokes.
Although there were no infarcts in the hippocampus, dysfunction in the CA1 subfield only occurred in patients who had infarcts in vascular territory that also supplied the hippocampal formation.
This was confirmed by comparing anatomical patterns across patients with infarcts in different vascular areas.
The finding may have therapeutic implications, the researchers said.
"Specifically, because excitotoxicity is thought to mediate CA1 dysfunction, our results suggest that as in the case of bilateral carotid occlusion, glutamate blocking agents such as memantine [Namenda] may be beneficial in patients who suffer an acute focal stroke in the hippocampal vascular territory," they said.
The brain in the age of old: The hippocampal formation is targeted differentially by diseases of late life
Annals of Neurology
Volume 64 Issue 6, Pages 698 - 706
Published Online: 23 Dec 2008
William Wu, BA 1, Adam M. Brickman, PhD 1, Jose Luchsinger, MD 1, Peter Ferrazzano, MD 1, Paola Pichiule, PhD 2, Mitsuhiro Yoshita, PhD 3, Truman Brown, PhD 1, Charles DeCarli, MD 3, Carol A. Barnes, PhD 4, Richard Mayeux, MD 1, Susan J. Vannucci, PhD 2, Scott A. Small, MD 1 *
1Taub Institute for Research on Alzheimer's Disease and the Aging Brain, Columbia University College of Physicians and Surgeons
2Department of Neuroscience in Pediatrics/Newborn Medicine, Weill Cornell Medical College, New York, NY
3Imaging of Dementia and Aging Laboratory, University of California at Davis, Sacramento, CA
4Evelyn F. McKnight Brain Institute, University of Arizona, Tucson, AZ
Abstract
Objective
To rely on the anatomical organization of the hippocampal formation in understanding whether and how late-life diseases such as diabetes and stroke contribute to age-related cognitive decline.
Methods
Magnetic resonance imaging (MRI) was used to document brain infarcts and to generate high-resolution functional maps of the hippocampal formation in 240 community-based nondemented elders (mean age, 79.7 years) who received a comprehensive medical evaluation. Sixty participants had type 2 diabetes mellitus, whereas 74 had MRI-documented brain infarcts, and the first analysis was designed to pinpoint hippocampal subregions differentially linked to each disorder. Then, guided by the results, additional functional MRI studies in aging rhesus monkeys and mice were used to test proposed mechanisms of dysfunction.
Results
Although both diabetes and brain infarcts were associated with hippocampal dysfunction, each was linked to separate hippocampal subregions, suggesting distinct underlying mechanisms. The hippocampal subregion linked to diabetes implicated blood glucose as a pathogenic mechanism, a hypothesis confirmed by imaging aging rhesus monkeys and a mouse model of diabetes. The hippocampal subregion linked to infarcts suggested transient hypoperfusion as a pathogenic mechanism, a hypothesis provisionally confirmed by comparing anatomical patterns across subjects with infarcts in different vascular territories.
Interpretation
Taken together with previous findings, these results clarify how diseases of late life differentially target the hippocampal formation, identify elevations in blood glucose as a contributing cause of age-related memory decline, and suggest specific interventions that can preserve cognitive health.
With increasing longevity, decreasing morbidity, and as older individuals expect to live intellectually challenging lives, cognitive aging has emerged as a major societal problem. Aging does not cause diffuse brain dysfunction but rather targets select brain areas, in particular, the frontal lobes and the hippocampal formation.[1-3]
The hippocampal formation itself, a structure vital for memory,[4] is made up of separate but interconnected subregions (Fig 1), and a wide range of studies have established that hippocampal subregions are differentially vulnerable to mechanisms of disease (as reviewed in Small's article[2]). Each subregion houses a molecularly distinct population of neurons,[5] providing a molecular basis for the observed vulnerability.[2] Therefore, as its basic anatomical unit of disease, evaluating hippocampal dysfunction at the level of the subregion can be helpful in clarifying causative ambiguities and can offer clues into mechanisms of dysfunction.
Indeed, although early stages of Alzheimer's disease (AD) undoubtedly contribute to age-related hippocampal dysfunction,[6-9] previous studies suggest that the hippocampal formation is also vulnerable to other diseases that occur commonly in later life, in particular, type 2 diabetes mellitus[10] and brain infarcts.[11] Nevertheless, because of the interaction and coexistence of late-life diseases, understanding how each disorder affects the hippocampal formation is challenging. Not only are diabetes and infarcts typically comorbid, but studies suggest that both interact with AD itself.[12][13] Notably, the early stages of AD occur years before clinical detection[14]; thus, hippocampal dysfunction observed in diabetes or infarcts may simply reflect an acceleration of AD pathophysiology. Because different disease mechanisms have been shown to target different subregions of the hippocampal formation, assessing the functional integrity of individual subregions in an aging and clinically characterized population of humans can be useful in resolving these issues.
With this goal in mind, we investigated a recently completed community-based study of nondemented elders who, together with a detailed medical assessment, were evaluated with magnetic resonance imaging (MRI) to document infarcts throughout the brain and to generate basal cerebral blood volume (CBV) maps of the hippocampal formation. As with other functional imaging variables, CBV is hemodynamically coupled to oxygen metabolism[15] and is tightly correlated with other basal measures of brain function, such as cerebral blood flow as measured with MRI or glucose uptake as measured with positron emission tomography.[16] Among all functional imaging approaches, measuring steady-state basal CBV with MRI provides the highest spatial resolution,[17] a feature that is important for visualizing individual hippocampal subregions. An additional advantage of CBV mapping is that it can be now used to investigate the hippocampal formation of animal models, both in nonhuman primates[18] and in rodents,[19] so that patterns observed in humans can be confirmed in experimental settings. Indeed, previous studies have used CBV mapping in humans and animal models to pinpoint dysfunction in select regions of the hippocampal formation in AD and normal aging.[18][19]
By measuring both infarcts and type 2 diabetes in a large dataset, we were able to first differentially link each disease to specific hippocampal subregions. Next, we tested hypotheses that emerged about underlying mechanisms by performing additional studies in nonhuman primates and a mouse model of diabetes, and by completing additional analyses on the human dataset. More than just linking diabetes and infarcts to hippocampal dysfunction, the series of reported findings clarifies general mechanisms underlying cognitive aging and suggests specific interventions to preserve cognitive health.
Discussion
By investigating the aging hippocampal formation in a large clinically characterized cohort, our results clarify causative factors contributing to cognitive aging, suggest possible mechanisms, and have potential therapeutic implications.
Consistent with the observation that the hippocampal subregions are differentially vulnerable to disease mechanisms,[2] the results highlight the importance of evaluating the hippocampal formation in a manner compatible with its anatomical complexity. As indicated by the multivariate results, both diabetes and infarcts were associated with global hippocampal dysfunction; it was only by examining the univariate results subregion by subregion that differences between the diseases were observed. Showing that each disease is differentially linked to separate subregions, and interpreting the findings in the context of previous studies, suggests distinct mechanisms of pathogenesis. Moreover, specific hypotheses could be generated and tested by pinpointing the individual subregions linked to each disease.
In the case of diabetes, our primary human findings suggested that the dentate gyrus is differentially vulnerable to blood glucose levels. Illustrating the utility of cross-species CBV mapping, we were then able to test this hypothesis, first in aging rhesus monkeys in which there is greater experimental control, and then, more definitively, in a mouse model of hyperglycemia. Although each study had it own limitations, as a composite, the human, monkey, and rodent findings establish that blood glucose differentially targets the dentate gyrus.
In the case of vascular disease, our results suggested that infarcts outside of the hippocampal formation are differentially linked to dysfunction in the CA1 subfield and the subiculum. Guided by previous studies,[11][26] we postulated that the link between CA1 dysfunction and infarcts might reflect transient hippocampal hypoperfusion. We would like to emphasize that numerous studies have established a link between hypoperfusion/ischemia and the CA1 subregion typically by transiently occluding large vessels such as the carotid[27][28] or middle cerebral artery[29][30] in rodent models, or by observational postmortem studies in patients who suffered a transient decrease in cardiac output.[31] Our findings extend these observations by suggesting that small or deep vessel infarcts can also indirectly target the CA1 subregion, as long as they occur within the hippocampal vascular territory. Nonhuman primates do not naturally develop this type of infarct, and experimentally causing an infarct selectively in the hippocampal vascular territory is difficult. Therefore, unable to test this hypothesis in animal models, we returned to the human dataset. This proposed mechanism was provisionally confirmed by showing that CA1 dysfunction was differentially observed only in subjects with infarcts in the hippocampal vascular territory. More direct confirmation will be provided by generating hippocampal CBV maps during the acute phases of stroke in different vascular territories.
In contrast with the CA1 subfield, we also found stroke-related dysfunction in the subiculum, and by showing that this effect occurs independent of vascular territory, our results suggest that there are additional mechanisms that link infarcts to hippocampal dysfunction. Nevertheless, strokes in the hippocampal vascular territory are common, and the proposed mechanism for CA1 dysfunction has potential therapeutic implications. Indeed, although previous postmortem studies have linked CA1 defects to vascular events, the ability to use CBV mapping to detect CA1 dysfunction in living subjects lends itself to in vivo detection and therapeutic monitoring. Specifically, because excitotoxicity is thought be mediate CA1 dysfunction, our results suggest that as in the case of bilateral carotid occlusion,[32] glutamate blocking agents such as memantine may be beneficial in patients who suffer an acute focal stroke in the hippocampal vascular territory.
A range of previous studies have established that the entorhinal cortex is differentially vulnerable to the early stages of AD,[6][33-35] which has been captured by hippocampal CBV imaging.[19] Because diabetes and infarcts may interact with AD pathology, it is theoretically possible that hippocampal dysfunction observed in stroke and diabetes is simply a reflection of these processes accelerating underlying AD pathology. By targeting other hippocampal subregions, our results suggest that both diabetes and infarcts can cause hippocampal dysfunction independent of AD pathophysiology. At the same time, our last analysis, showing that the entorhinal cortex is differentially sensitive to insulin in a stroke-dependent manner, provides an interesting anatomical site of convergence linking AD, diabetes, and infarcts. Because this final analysis was based on a relatively low number of subjects, however, further studies are required to better understand this complex relationship.
Showing that blood glucose selectively targets the dentate gyrus is not only our most conclusive finding, but it is the one most important for normal aging, that is, hippocampal dysfunction that occurs in the absence of disease states, such as AD, infarcts, and diabetes. Indeed, cognitive studies have established that normal age-related hippocampal dysfunction begins quite early,[36] typically during the fourth decade of life, before the onset of age-related diseases. Furthermore, age-related hippocampal dysfunction occurs in all nonhuman mammals,[37] which do not typically experience development of AD, stroke, or diabetes. Consistent with this, our cross-species findings document that the detrimental effects of glucose on the hippocampus[23][24] occur independent of AD and infarcts, and our monkey findings in particular suggest that it occurs independent of overt diabetes.
With increasing longevity and decreasing morbidity, age-related hippocampal dysfunction has emerged as a cognitive epidemic. Nevertheless, underlying causes of age-related hippocampal dysfunction have remained unknown. Importantly, converging evidence in humans,[38][39] nonhuman primates,[18][40] and rodents[18][19] has suggested that the dentate gyrus is differentially vulnerable to normal aging.[41] At the same time, an independent series of cross-species studies has shown that glucose regulation worsens with advancing age.[42][43] Taken together, our findings suggest that glucose dysregulation is as at least one systemic cause underlying age-related hippocampal dysfunction.
Beyond the obvious conclusion that preventing late-life disease would benefit the aging hippocampal formation, our findings suggest that maintaining glucose control, even in the absence of disease, should be strongly recommended to preserve cognitive health. More specifically, our findings predict that any intervention that causes a decrease in blood glucose should increase dentate gyrus function and would, therefore, be cognitively beneficial. In fact, separate studies examining the effects of physical exercise support this prediction. Imaging studies in humans and mice have documented that, among all hippocampal subregions, physical exercise causes a differential improvement in dentate gyrus function.[44] By improving glucose metabolism, physical exercise also reduces blood glucose.[45] It is possible, therefore, that the cognitive-enhancing effects of physical exercise are mediated by the beneficial effect of lower glucose on the dentate gyrus. Whether through physical exercise or other behavioral or pharmacological interventions, our results suggest that improving glucose metabolism is a clinically tractable approach for ameliorating the cognitive slide that occurs in all of us as we age.
Results
Diabetes and Brain Infarcts Are Differentially Linked to Hippocampal Dysfunction
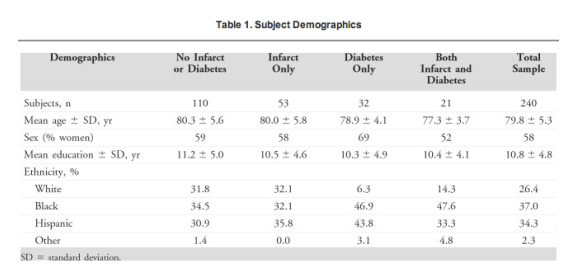
Among the 240 human subjects who participated in this study, 60 subjects had type 2 diabetes. A total of 216 subjects were also evaluated for vascular disease, and 74 subjects had documented brain infarcts. When comparing demographics (Table 1), we found the diabetes and infarct groups to be similar in age [F (3, 216) = 2.221; p = 0.087], sex distribution [X2(3) = 1.606; p = 0.658], education [F (3, 214) = 0.430; p = 0.732], and ethnicity [2(9) = 13.158, p = 0.156], although those with diabetes were more likely to be black.
Although subjects with diabetes had relatively more infarcts than those without diabetes; this difference was small and not statistically significant (X2 = 0.34; p = 0.40; Fig 2A). Therefore, in our first series of analyses, we independently investigated the relation between hippocampal dysfunction and diabetes, and between hippocampal dysfunction and infarcts. To identify hippocampal subregions linked to each disorder, we constructed an analysis of variance (ANOVA) model in which CBV measured in the hippocampal subregions was included as the dependent variables, group (disease vs no disease) was included as the fixed factor, and age, sex, ethnicity, and education were included as covariates. Results showed that diabetes was associated with global hippocampal dysfunction, as indicated by the model's multivariate test (F = 5.88; p = 0.0002), and univariate tests showed that this effect was selectively driven by dysfunction in the entorhinal cortex (F = 4.4; p = 0.036) and the dentate gyrus (F = 10.7; p = 0.001) (see Figs 1C and 2B). In contrast, although infarcts were also associated with global hippocampal dysfunction (F = 2.92; p = 0.022), the effect was selectively driven by dysfunction in the CA1 subfield (F = 3.97; p = 0.047) and the subiculum (F = 7.38; p = 0.007) (see Figs 1C and 2C).
Next, to test for a potential relation between the two diseases, we repeated the ANOVA, but both diabetes and infarcts were included as covariates in a single model. Replicating the first series of studies, each disease was selectively associated with global hippocampal dysfunction (diabetes: F = 5.41; p = 0.0004; infarcts: F = 2.85; p = 0.025). However, although in the case of infarcts the effect was driven by the same hippocampal subregions (CA1 subfield: F = 4.48; p = 0.036; subiculum: F = 7.24; p = 0.008), in the case of diabetes, the effect was driven selectively by the dentate gyrus (F = 14.27; p = 0.0002), not the entorhinal cortex (F = 0.79; p = 0.38).
Finally, in post hoc analyses, we further examined hippocampal effects by comparing subjects with infarcts only versus those without diabetes or infarcts, and we examined hippocampal effects by comparing subjects with diabetes only versus those without diabetes or infarcts. The results again showed that subjects with infarcts have greatest dysfunction in the CA1 subfield (F = 3.70; p = 0.042) and the subiculum (F = 6.41; p = 0.012), whereas subjects with diabetes have selective dysfunction in the dentate gyrus (F = 10.59; p = 0.001).
These results suggest that the dentate gyrus is selectively and differentially linked to diabetes, that the CA1 subfield and the subiculum are selectively and differentially linked to infarcts, and that there is complex interplay linking the entorhinal cortex to diabetes and infarcts. The following sections further investigate each of these three observations separately.
Relation between Diabetes and the Aging Hippocampal Formation
Type 2 diabetes is associated with increases in blood insulin and glucose levels, and impaired glucose regulation in particular has been found to affect hippocampal function.[23][24] To determine whether blood glucose or insulin might underlie dentate gyrus dysfunction, we investigated their relation with hippocampal CBV. Levels of blood glucose and insulin were measured in a subset of 181 subjects. A partial correlation analysis was performed between these measurements and hippocampal CBV, covarying for age, sex, ethnicity, and education. Importantly, because some subjects had fasting blood measurements whereas others did not, this factor was also included as a covariate. Results showed that among all hippocampal subregions, blood glucose levels were inversely and selectively correlated with dentate gyrus CBV (beta = -0.163; p = 0.032) (Table 2). In addition, blood glucose levels were found to selectively and inversely correlate with total recall on the Selective Reminding Test (beta = -0.180; p = 0.018), a cognitive measure of hippocampal function.[25] Using the same statistical model, we found that blood insulin levels did not correlate with hippocampal CBV, although we observed a trend for an inverse relation with the entorhinal cortex (beta = -0.128; p = 0.08) (see Table 2).
These findings suggest that increases of blood glucose may differentially target the dentate gyrus. Nevertheless, these results are limited by the fact that only some subjects had fasting glucose measurements and by the general limitation of human-based studies in which it is difficult to control for all the factors that can potentially affect CBV. To test the hypothesis that increases in blood glucose level is differentially linked to the dentate gyrus in a more controlled experimental setting, we turned to animal models. We returned to a previous study in which we mapped hippocampal CBV in aging rhesus monkeys.[18] Among the 14 subjects imaged, 11 had blood glucose levels measured within a few months of the imaging study. A correlation analysis demonstrated that among the hippocampal subregions, blood glucose levels were selectively and inversely correlated with dentate gyrus CBV (beta = -0.64; p = 0.033) (Fig 3A).
Although the findings in rhesus monkeys support the interpretation that blood glucose differentially targets the dentate gyrus, they also have limitations. First, insulin levels were not measured; therefore, we cannot exclude that it might have contributed to the finding. Second, and more importantly, the findings in monkeys were only correlational.
To address both limitations, we turned to a streptozosin mouse model of diabetes. By killing insulin-producing cells of the pancreas, administration of streptozosin causes an abnormal increase in blood glucose level by reducing insulin. Streptozosin was administered to 3-month-old mice, and at 6 months of age, CBV maps of the hippocampal formation were generated in four treated mice and four nontreated mice. An ANOVA was performed, in which CBVs measured from the entorhinal cortex, dentate gyrus, CA1, and subiculum were included as the dependent variables, and group (treated vs nontreated) was included as the fixed factor. Results showed that, compared to controls, streptozosin-treated mice had dominated dysfunction in the dentate gyrus CBV (F = 6.62; p = 0.042) (see Fig 3B).
Relation between Brain Infarcts and the Aging Hippocampal Formation
None of the subjects had infarcts in the hippocampal formation, according the criteria described earlier (see Subjects and Methods). Nevertheless, an ANOVA, covarying for age, sex, ethnicity, and education, showed that compared with subjects without infarcts, those with infarcts in regions other than the hippocampal formation had poorer performance on the Selective Reminding Test (F = 9.59; p = 0.002).
The observation that CA1 dysfunction was linked to infarcts, as documented earlier, suggested a possible mechanism for how strokes in extrahippocampal sites might cause hippocampal dysfunction. Specifically, previous studies have shown that the CA1 subfield is differentially vulnerable to transient hypoperfusion.[11] Furthermore, studies have suggested that during the evolution of an infarct in a particular brain region, transient hypoperfusion occurs throughout the vascular territory supplying that region.[26] Taken together, we postulated that CA1 dysfunction may occur only when infarcts occur in a vascular territory that also supplies the hippocampal formation, namely, the anterior choroidal and posterior cortical arteries. To test this hypothesis, we subdivided the 74 subjects with stroke into 3 groups: (1) 22 subjects who had infarcts in a vascular territory that supplies the hippocampal formation (basal ganglia, thalamus, internal capsule, and occipital cortex), (2) 52 subjects who had infarcts in other vascular territories, and (3) 142 subjects who were infarct free. Confirming the hypothesis, we observed CA1 dysfunction only in subjects with infarcts in the hippocampal vascular territory (F = 7.41; p = 0.007) (Fig 4).
Compared with subjects without infarcts, subiculum dysfunction was observed in subjects with infarcts in the hippocampal territory (F = 6.56; p = 0.035), and subiculum dysfunction was observed in subjects with infarcts in other territories (F = 4.11; p = 0.044) (see Fig 4). This suggests that stroke-associated dysfunction in the subiculum is linked to other mechanisms.
Interrelation between Diabetes and Brain Infarcts
As reported earlier, the link between diabetes and entorhinal cortex CBV was found to be stroke dependent; furthermore, there was a suggestion that entorhinal cortex CBV may be correlated with insulin levels. We postulated that the entorhinal cortex may be selectively associated with insulin levels only among subjects with infarcts in the hippocampal vascular territory.
To test this hypothesis, we repeated the correlation analysis among the three stroke subgroups and found an inverse relation between entorhinal cortex CBV and blood levels of insulin only among the subjects with infarcts in the hippocampal vascular territory (beta = -0.58; p = 0.027) (Fig 5). No correlations was observed among subjects with infarct in other territories (beta = -0.06; p = 0.78), or among subjects without infarct (beta = -0.04; p = 0.69). Of note, only one of the subjects with stroke in the hippocampal vascular territory had a diagnosis of diabetes.
Subjects and Methods
Subjects
HUMANS.
Participants were part of a community-based study of elderly subjects (65 years and older) who, together with brain imaging,[20] received a detailed neuropsychological, neurological, and medical evaluation.[21] Within this cohort, hippocampal CBV maps were generated in 240 subjects. Relying on clinical criteria, a consensus conference made up of neurologists and neuropsychologists determined that all subjects were nondemented.
Type 2 diabetes mellitus was ascertained by self-report or by the use of diabetes medications. Brain infarcts were ascertained by MRI, relying on whole-brain fluid-attenuated inverse recovery T2-weighted sequence (TR = 11,000 milliseconds; TE = 144 milliseconds; 3mm slice thickness) and T1-weighted high-resolution anatomical sequence (TR = 20 milliseconds; TE = 2.1 milliseconds; 1.3mm slice thickness) acquired in the axial orientation.[22] Lesions greater than or equal to 3mm were considered for classification as infarct.
Serum glucose and insulin levels were measured in micro-international units per milliliter (IU/ml) from serum collected within 1 month of the brain MRI and frozen at -70·C. Glucose levels were measured on a Hitachi automated spectrophotometer (model 704; Hitachi Limited, Tokyo, Japan) using commercial kits obtained from Wako Chemicals (Richmond, VA). Insulin levels were measured using a solid-phase chemiluminescent enzyme immunoassay (Immulite; Diagnostic Products, Los Angeles, CA). The intraassay coefficient of variation was 4.7%, and the interassay coefficient of variation was 8.2%. The reference insulin range for this assay is 6 to 27IU/ml. The study was approved by the appropriate institutional ethics review committee, and all subjects gave written informed consent to participate.
NONHUMAN PRIMATES.
Rhesus monkeys (Macaca mulatta) 7 to 31 years of age were used as subjects in this experiment. The animal colony rooms were on a 12-/12-hour light/dark (6 AM to 6 PM light) schedule, maintained at 26·C, and all animals were trained to deliver their arms so that morning fasting blood samples could be obtained. The arm was shaved, wiped with alcohol, and approximately 1ml blood was drawn from a vein in the arm. The blood was centrifuged for 10 minutes at 3,500 rpm, cooled, and glucose levels determined using a standard serum chemistry panel in the Veterinary Medicine Teaching Hospital's clinical laboratory at the Primate Center in Davis. The test was performed using a Hitachi 917 (Roche Biomedical, Indianapolis, IN) instrument. All protocols were approved by the University of California Davis Animal Care and Use Committee.
MICE.
Male C57/Bl6 mice were purchased from Jackson Laboratories (Bar Harbor, ME) at 4 weeks of age. At 5 weeks of age, diabetes was induced by 5 daily injections (intraperitoneally) of streptozocin (55mg per kilogram body weight) freshly dissolved in 0.2M citrate buffer, pH 4.5. Control animals received an equal volume of diluent. Blood glucose was analyzed in tail blood with a glucometer at 5 days after the last injection, and diabetes was confirmed by a blood glucose value 250mg/dl. All protocols were approved by the Columbia University Animal Care and Use Committee.
Functional Imaging
HUMANS.
The technical and analytic details of how CBV maps of the human hippocampal formation were generated has been described previously.[19] In brief, a 1.5-Tesla scanner (Philips Intera, Eindhoven, The Netherlands) was used to acquire oblique coronal three-dimensional T1-weighted images (TR = 20 milliseconds; TE = 6 milliseconds; flip angle = 25 degrees; in-plane resolution = 0.86 _ 0.86mm; slice thickness = 3mm), perpendicular to the hippocampal long axis (see Fig 1A), before and 4 minutes after intravenous administration of gadolinium-pentate (Omniscan, 0.1mmol/kg). Then, the postcontrast images were subtracted from precontrast images, and the difference in the superior sagittal sinus, which serves as an estimate of the image intensity change of 100% blood, was recorded.[17][19] Finally, the subtracted image was divided by the difference in the top four pixels measured from the sagittal sinus and multiplied by 100, yielding percentage CBV.
A single ideal slice was always identified, anterior to the lateral geniculate nucleus and posterior to the uncus, that contains all hippocampal subregions and provides sufficient anatomical information to parse the subregions[19] (see Fig 1B). Because spatial coregistration across subjects is problematic when evaluating small regions in clinical populations, strict anatomical criteria[19] were used to identify the following regions of interest within the hippocampal formation: entorhinal cortex, dentate gyrus, CA1 subfield, and subiculum (see Fig 1B). On an individual basis, mean CBV values were measured for each hippocampal region of interest and used for group data analysis.
NONHUMAN PRIMATES.
As described previously,[18] CBV maps of the monkey hippocampal formation were generated in a 1.5-Tesla scanner using a gadolinium-based approach nearly identical to the method used in humans. The main difference was that T1-weighted images (TR = 50 milliseconds; TE = 5 milliseconds; flip angle = 35 degrees; in-plane resolution = 0.62 _ 0.62mm; slice thickness = 2mm) and hippocampal parsing were optimized for the monkey brain.
MICE.
The technical and analytic details of how CBV maps of the mouse hippocampal formation were generated has been described previously.[19] In brief, a 9.4-Tesla scanner (Bruker, AVANCE 400WB, Bilerica, MA) was used to acquire axial T2-weighted images (TR/TE = 2,000/70 milliseconds; field of view = 19.6mm; acquisition matrix = 256 _ 256; 8 slices; slice thickness = 0.6mm), perpendicular to the hippocampal long axis, before and 45 minutes after intraperitoneal administration of gadolinium-pentate (Omniscan, 013mmol/kg). Isofluorane was chosen as the anesthetic agent because of its minimal effects on hemodynamic coupling, and heart rate, respiratory rate, and SaO2 were monitored throughout. Relative CBV maps were generated according to this formula: CBV R2 = ln (Spre/Spost)/TE, where TE = effective echo time, Spre = signal before contrast, and Spost = signal contrast agent reaches steady state. The derived image was then divided by the maximum four-pixel signal value of the posterior cerebral vein to control for differences in levels of contrast yielding relative CBV.
As in humans and monkeys, an ideal single slice, acquired through the body of hippocampal formation, was identified that contains all hippocampal subregions and has sufficient anatomical information to parse the subregions.[19] Strict anatomical criteria[19] were used to identify the following regions of interest within the hippocampal formation: entorhinal cortex, dentate gyrus, CA1 subfield, and subiculum. On an individual basis, mean CBV values were measured for each hippocampal region of interest and used for group data analysis.
|
|
| |
| |
|
|
|