| |
The cell biology of bone metabolism
|
| |
| |
Journal of Clinical Pathology 2008;61:577-587; doi:10.1136/jcp.2007.048868
H K Datta1, W F Ng1, J A Walker2, S P Tuck3, S S Varanasi1
1 Institute of Cellular Medicine, Musculoskeletal Research Group, The Medical School, University of Newcastle, Newcastle upon Tyne, UK
2 Department of Pathology, Leeds General Infirmary, Leeds, UK
3 James Cook University Hospital, Middlesbrough, UK
"Recent evidence shows that osteoblast function may contribute to obesity and glucose intolerance. This suggests that the skeleton exerts an endocrine regulation of energy metabolism and thereby may contribute to the onset and severity of metabolic disorders"
"the interaction between the immune system and bone cells is of particular interest, leading to the emergence of a new discipline termed osteoimmunology. The role of lymphocytes and a number of key cytokines in the regulation of osteoclastogenesis and osteoblast function..."....."In young adulthood, formation and resorption are in approximate balance, but with ageing there is a net loss of bone".....In order for the strength of the bone to be maintained, the process of bone turnover must be carefully regulated....[from Jules: peak bone mass occurs for people in their early 20s, and this process can be interrupted by a chronic viral infection such as HIV. We have not studied the effect of HIV-infection during these years on bone metabolism and how it might lead to the high rates of bone loss we see in HIV-infected individuals]"
"The basic multicellular unit of bone comprises the osteocytes, osteoclasts and osteoblasts. Its activity of the unit is regulated by mechanical forces, bone cell turnover, hormones (e.g. parathyroid hormone (PTH), growth hormone (GH)), cytokines and local factors"
"It has long been recognised that mechanical stress induced by weight-bearing exercise increases osteoblast activity. Indeed, the absence of mechanical stimulation resulting from prolonged immobilisation or microgravity causes severe bone loss....Many cytokines are important regulators of osteoclast and osteoblast function, and a majority of these cytokines are produced by cells of the immune system.....most act indirectly by regulating the interactions between osteoclasts and osteoblasts.......pro-inflammatory cytokines, such as IL-1, TNF-{alpha}, and IL-6, appear to act synergistically on osteoclastogenesis and promote osteoclast function....[from Jules: chronic disease or virus-infection can disregulate cytokines and be associated with bone loss]"
T cells
T cells rapidly up-regulate surface expression of RANKL on activation, and are therefore capable of regulating osteoclast differentiation and activity. Indeed, activated T cells have been shown to promote osteoclastogenesis in vitro, mediate bone erosion in inflammatory joint disease and play an essential role in the pathogenesis of osteoporosis in ovariectomised mice.144-146 In contrast, resting T cells actively suppress osteoclastogenesis in a cell:cell contact independent manner.147 RANKL is also expressed on memory T cells, but not on naive T cells, but the physiological significance of such differential expression is unclear.148 Apart from RANKL, T cells produce many other cytokines that modulate osteoclast activity and osteoclastogenesis; TNF-{alpha}, IL-6 and IL-17 are stimulatory, while IL-4, IL-13 and IL-10 are inhibitory.149 Since the range of cytokines produced by different T cell subsets show considerable variability, so does their effect on osteoclast number and activity. In addition to their effect on osteoclast function, T cells may also interact with osteoblasts directly and exert effects on bone turnover indirectly via their interactions with dendritic and B cells.150 151
Osteoimmunology
Recent efforts are beginning to define the nature of the interactions between the immune and skeletal systems (fig 6). The realisation of the intricate nature of this interaction has been supported by the fact that a number of cell surface receptors, cytokines and signalling pathways serve a critical role both in the immune and skeletal systems. The close interaction between immune progenitors and skeleton is facilitated by their proximity in the bone marrow. Therefore, osteoimmunology, a discipline that encompasses the complex interactions and overlapping functions of the immune system and bone cells has emerged over recent years.
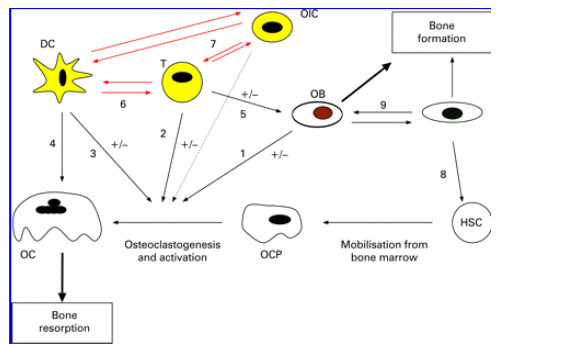
Figure 6 Interactions between the immune system and bone. OB play a key role in regulating osteoclastogenesis and activation of OC (1). T cells also regulate these processes both directly via RANKL-RANK interaction and production of cytokines (2), and indirectly via their effects on OB (5) and DC (6). The overall effects may be stimulatory or inhibitory, depending on the T cell subsets and the cytokines produced as well as other local factors. DC produce cytokines that can promote or suppress osteoclastogenesis (3); DC also interact with T cells (6). It has also been suggested that DC can be transformed to become OC in inflammatory milieu (4). Other immune cell types may affect OC function by interaction with T cells and DC (7). The direct effects of other immune cell types in osteoclastogenesis and OC activation have not been extensively investigated (dotted arrow). Finally, osteocytes may play a role in regulating haemopoiesis (8) and regulate osteoblast activity (9). OIC, other immune cells; T, T cells; DC, dendritic cells; OC, osteoclasts; OB, osteoblasts; OCP, osteoclast precursors; HSC, haemopoietic stem cells.
Cytokines and growth factors
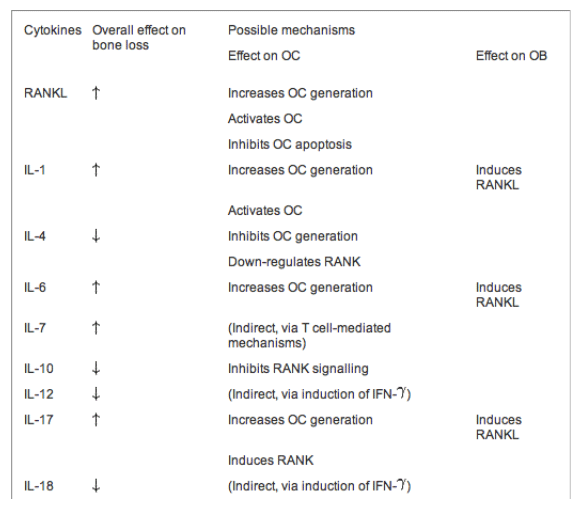
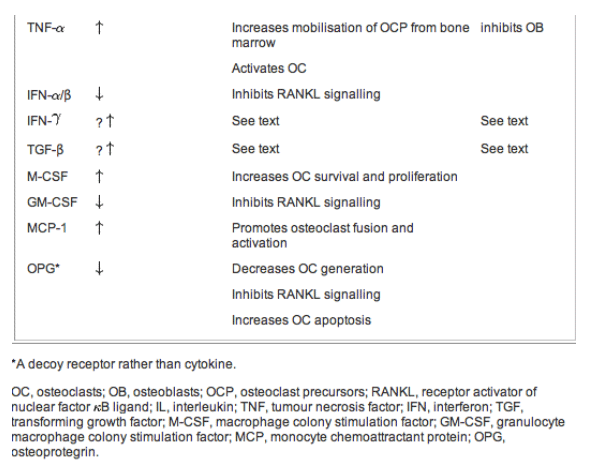
Many cytokines are important regulators of osteoclast and osteoblast function, and a majority of these cytokines are produced by cells of the immune system.156 The precise mechanisms of how these cytokines regulate osteoblast and osteoclast activities have not been fully defined; most act indirectly by regulating the interactions between osteoclasts and osteoblasts. For instance, many cytokines have been shown to regulate the expression of RANK and RANKL on osteoclast and osteoblasts respectively, or modulate the intracellular signalling mediated by RANK. In general, pro-inflammatory cytokines, such as IL-1, TNF-{alpha}, and IL-6, appear to act synergistically on osteoclastogenesis and promote osteoclast function.157-162 Table 1 summarises the effects of the various cytokines on bone metabolism and the possible underlying mechanism(s) of action.
TNF-{alpha} enhances RANKL-mediated stimulation of osteoclasts by activating NF-{kappa}B.158 TNF-{alpha} also enhances the mobilisation of osteoclastogenesis progenitors from bone marrow and inhibits osteoblastic activity.159 Like TNF-{alpha}, IL-1 activates NF-{kappa}B, which promotes osteoclast survival and induces RANKL expression on osteoblasts.160 There are two forms of IL-1, IL-1{alpha} and IL-1β, but their biological activities are indistinguishable; cells of monocytic lineage on activation predominantly produce IL-1β, while IL-1{alpha} is released by dying cells and is not normally found in the circulation.161 Osteoblasts and stromal cells are the main source of IL-6 in bone; the effect of IL-6 on bone resorption is mediated primarily via stimulation of osteoclasts.162 163 IL-17, a key cytokine that mediates autoimmune inflammatory arthritis, also augments osteoclastogenesis.164 In contrast, Th2 cytokines such as IL-4, IL-13 and IL-10 suppress osteoclastogenesis and osteoclast activation.165 Interestingly, a recent report suggests that IL-4 may play an important role in CD4+CD25+Foxp3+ regulatory T cell-mediated suppression of osteoclastogenesis.166 Another class of cytokines that inhibits osteoclastogenesis is the interferons. IFN-β suppresses RANKL-induced expression of c-Fos; IFNβ-/- mice have increased osteoclast activity and are osteoporotic.167
Table 1 Effects of various cytokines produced by the immune system on bone metabolism
ABSTRACT
Contrary to the commonly held misconception, bone is a relatively dynamic organ that undergoes significant turnover as compared to other organs in the body. This review details how complex intercellular signalling, between the osteoprogenitor cells and mature osteoblasts, osteocytes and osteoclasts, regulates and balances activities of bone cells during remodelling and growth. Both systemic, as well as local autocrine and paracrine factors are discussed. A number of recent important advances in cell biology of bone have led to a new paradigm in understanding of the subject. In this regard, the interaction between the immune system and bone cells is of particular interest, leading to the emergence of a new discipline termed osteoimmunology. The role of lymphocytes and a number of key cytokines in the regulation of osteoclastogenesis and osteoblast function is critically examined. The intracellular signalling regulating key cellular pathways involved in cell differentiation and activity are outlined. The emerging evidence of osteocytes as mechanosensors as well as regulators of mineralisation is discussed.
Bone is a specialised connective tissue hardened by mineralisation with calcium phosphate in the form hydroxyapatite ([Ca3(PO4)2]Ca(OH)2). Bone has well recognised mechanical functions: it provides rigidity and shape, protection and support for body structures, and aids locomotion. Contrary to popular belief, bone is in fact a highly dynamic structure undergoing constant remodelling. This bone turnover allows the bone to repair itself, for example after fracture, and to adapt to the forces placed on it. In childhood there is a very high rate of bone turnover in which formation exceeds resorption. In young adulthood, formation and resorption are in approximate balance, but with ageing there is a net loss of bone. The rate of bone turnover, collagen matrix, size, structure, geometry and density all combine to determine the bone's overall mechanical properties. Defects in these parameters will result in diseases such as osteoporosis, Paget's disease of bone, osteopetrosis and osteogenesis imperfecta.
In order for the strength of the bone to be maintained, the process of bone turnover must be carefully regulated. This review will discuss the systemic and local regulatory mechanisms, as well as the cell-cell interactions involved in bone development, morphogenesis and remodelling. The emerging role of osteocytes as a pivotal player in the control of osteoblast and osteoclast function will be described, along with recent insights into the role of the immune system in the regulation of bone turnover.
BONE STRUCTURE AND CELL TYPES
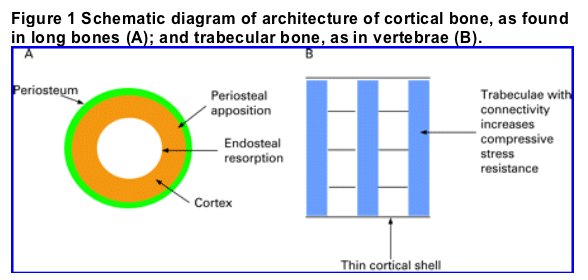
The skeleton is comprised of long bones, such as humerus, femur and tibia, and flat bones, such as skull, scapula and ileum. There are two main histological types of mature bone: cortical or compact bone which has a dense, ordered structure and cancellous or trabecular bone which is lighter, less compact and has an irregular structure. Cortical or compact bone is found mainly in the shaft of long bones and the surfaces of flat bones. It is composed of bone laid down concentrically around central canals known as Haversian systems. These contain blood vessels, lymphatics, nerves and connective tissue. A concentric layer of rings or lamellae of bone matrix surround each Haversian canal. Within the lamellae are tiny spaces called lacunae containing osteocytes. Trabecular or cancellous bone is less numerous and forms the ends of long bones and the inner parts of flat bones. It contains interconnecting plates and bars called trabeculae, with intervening marrow lending it a honeycomb appearance. The trabeculae are aligned along lines of stress; this connectivity adds considerably to its strength. The specific anatomical distribution of cortical and trabecular bone reflects their respective compressive and tensile strengths. Thus trabecular bone is ideally suited to withstanding compressive stress and hence is the predominant bone in vertebrae.
A further important element in the structure of bone is the collagen network. In adults, collagen fibres adopt a preferential orientation resulting in the formation of lamellar bone. In cortical bone the lamellae are concentrically arranged, and in trabecular bone they are parallel to one another. Woven bone is formed in the growing skeleton and in various pathological conditions in the adult. This type of bone has a random arrangement of collagen fibres, with non-uniformly sized and distributed osteocytes.
In general each bone has an outer layer of cortical bone overlying trabecular bone and the medullary cavity. The cortical bone has an outer membrane called the periosteum. The periosteum has two layers: an outer fibrous layer and an inner one, which has osteogenic potential and lays down new bone allowing the bone to enlarge, a process known as periosteal apposition. The inner surface of the cortex has another lining called the endosteum (fig 1). Bone tends to undergo resorption from the endosteal surface. Both the periosteum and endosteum contain osteoblasts and osteoclasts and their progenitors. Osteoblasts and osteoclasts function in a coordinated manner, by their respective bone forming and resorbing activities, to carry out remodelling, growth and repair.
THE CYCLE OF BONE TURNOVER
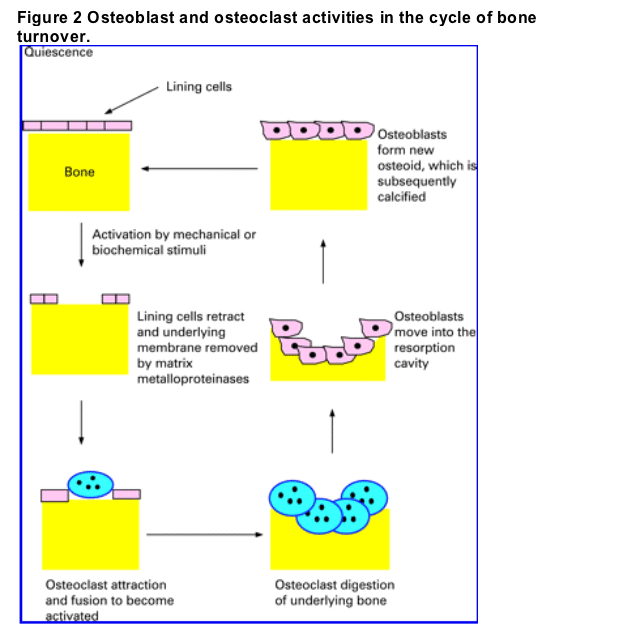
The basic multicellular unit of bone comprises the osteocytes, osteoclasts and osteoblasts. Its activity of the unit is regulated by mechanical forces, bone cell turnover, hormones (e.g. parathyroid hormone (PTH), growth hormone (GH)), cytokines and local factors. The activation process is at least partly regulated by the osteocytes, which detect mechanical stress and respond to biochemical stimuli. Activation results in the lining cells of the endosteal surface being retracted, and digestion by matrix metalloproteinases of the endosteal collagenous membrane. Osteoclasts are then recruited, followed by fusion of activated osteoclasts to become multinucleated osteoclasts. Activated osteoclasts mediate resorption of the underlying bone. Subsequently, osteoblasts are recruited to the resorption cavity and lay down new osteoid, which eventually becomes calcified; this process is completed in approximately 3-6 months (fig 2). The rate of bone turnover varies according to the type of bone, being highest in sites where trabecular bone predominates such as vertebrae and lowest at sites high in cortical bone such as the hip.
DIFFERENTIATION AND LOCAL REGULATION OF BONE CELLS
Osteoblasts
Considerable progress has been made in identifying morphogens, signalling pathways and transcriptional regulators mediating differentiation of mesenchymal stem cells into osteoprogenitors (fig 3). During skeletal development osteoblast differentiation and deposition of bone matrix occurs, and involves spatiotemporal coordination of interaction among diverse endocrine, paracrine and autocrine factors. Osteoblast differentiation is stimulated by several factors at different stages of differentiation. Thus, epidermal growth factor promotes renewal of mesenchymal stem cells; self renewal has been found to be critically dependent on the expression of certain genes, such as stem cell antigen 1 (Sca-1/Ly- 6A).1 2 Osteoblast stimulating facor (OSF-)-1, also known as pleiotrophin (PTN) or heparin-binding growth-associated molecule (HB-GAM), is chemotactic for the osteoprogenitor and stimulates mature osteoblast activity.3 4 Other factors such as PTH, GH, prostaglandins and insulin-like growth factor-1 (IGF-1) exhibit more pervasive effects by affecting renewal of mesenchymal stem cells (MSC) and stimulating osteogenic differentiation of bone morphogenetic protein (BMP)-primed cell populations.
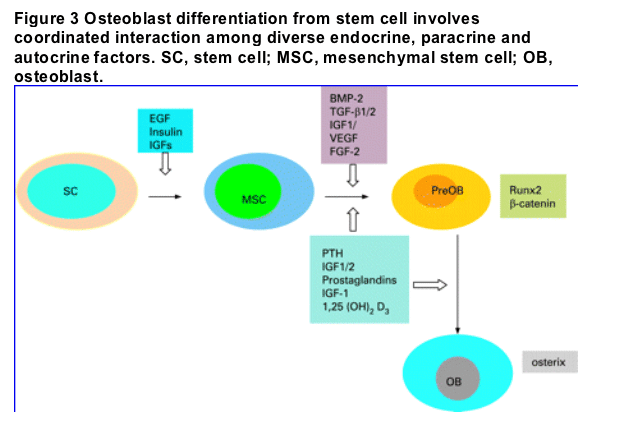
The development of the skeleton, like many other tissues, involves complex spatiotemporally regulated molecular interaction between Wnt/catenin and transforming growth factor-_/BMP mediated signalling pathways. The expression of Runx2, a key osteoblast differentiation transcription factor, is required for multipotent MSC to differentiate into osteoblastic lineage. Runx2 also inhibits the differentiation of MSC into adipocytic and chondrocytic lineages. BMP2 and WNT pathways jointly promote RUNX2 expression to promote osteoblast differentiation.
Canonical Wnt pathway
Wingless-ints (Wnts), secreted lipid-modified glycoproteins, activate three distinct intracellular signalling pathways, including the Wnt/β-catenin pathway, which is commonly referred to as the canonical Wnt pathway (fig 4).5-10 Genetic studies in human and animal models suggest that the canonical Wnt pathway, together with BMP signalling and key transcription factor RUNX2(CBFA1/AML3), has an important role in skeletal development, osteoblast differentiation and bone formation.11-19 A member of the lipoprotein receptor family, low-density lipoprotein receptor-related protein 5 (LRP5), plays a critical role in canonical Wnt signalling by forming a frizzled/LRP5/6 receptor complex, and Wnts activate the canonical pathway by interacting with receptors of the frizzled family and co-receptors of the LRP5/6 family.20 The importance of LRP5 has been shown by the observations that activating and inactivating mutations of the gene resulted in high bone mass and the osteoporosis-pseudoglioma syndrome respectively.21-23
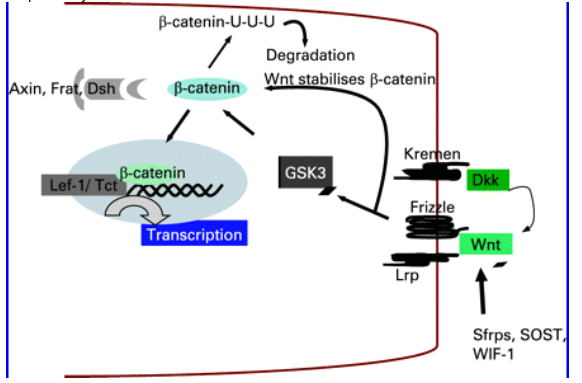
Figure 4 Canonical Wnt/catenin pathway in osteoblast differentiation. Wnt/β-catenin pathway signals through LRP-5 or LRP-6, leading to inhibition of glycogen synthase kinase (GSK)-3, thereby preventing phosphorylation and degradation of β-catenin. The accumulated β-catenin translocates to the nucleus and activates T cell-specific transcription factors. Wnts also activate non-canonical pathways that include the G-protein and dishevelled (Dsh) mediated c-Jun NH2-terminal kinase (JNK) signalling, but the significance of the non-canonical pathways is unclear. The extracellular secreted proteins (WIF-1 and Sfrps) inhibit by binding Wnt or by blocking LRP5/6 (Dkks or sclerostin).
Wnt signal transduction involves stabilisation of β-catenin by inhibiting GSK-3-mediated β-catenin phosphorylation. Unphosphorylated β-catenin accumulates, and translocates to the nucleus, binds LEF/TCF transcription factors and activates the transcription of downstream genes (fig 4).10 β-catenin-/- transgenic mice revealed an essential role of Wnt signalling in osteoblast differentiation.12 24 25 Dickkopf family members (Dkk1 and Dkk2) and secreted frizzled related proteins (Sfrps) are families of extracellular proteins that negatively modulate canonical Wnt signalling; Sfrps bind directly to Wnts and prevent their association with Lrp and Fzd receptors.26-35 Dkks limit the availability of Lrp5/6 receptors to Wnts by sequestering Lrp5/6 into complexes with the transmembrane protein Kremens (Kringle-containing protein marking the eye and the nose) and may promote its lysosomal destruction (fig 4); Kremens have high affinity for Dkk.36 37 Sclerostin, a SOST gene product, shares many characteristics with the Wnt antagonist dickkopf-1 in antagonising BMP-stimulated bone formation and BMP- and Wnt-induced Wnt activation. Sclerostin inhibits BMP-stimulated bone formation, but does not affect BMP signalling.38-45 Instead, it antagonises Wnt signalling in osteoblastic cells. High bone mass in sclerosteosis and van Buchem disease may, therefore, result from increased Wnt signalling.
In the absence of Wnt signalling, β-catenin is phosphorylated by glycogen synthase kinase 3 (GSK3), which promotes its ubiquitination (U) by E2 ligases and degradation by the proteosome.46 47 The scaffolding proteins (Axin, Frat and Dishevelled (Dsh)) mediate the degradation of phosphorylated β-catenin by creating docking sites for F-box protein/E2 ligase complexes.48-51 Wnt signalling prevents β-catenin degradation, β-catenin then enters the nucleus where it displaces corepressors (CoR) from T cell-factor/lymphoid-enhancer-factor (Tcf/Lef) and recruits transcriptional co-activators to stimulate expression of many genes; such as those involved in cycle progression (e.g. cyclin D1) and survival (e.g. c-myc) (an updated list of genes modulated by Wnt signalling is available on: http://www.stanford.edu/~rnusse/wntwindow.html).
Bone morphogenetic proteins
There are at least 30 bone morphogenetic proteins (BMPs) that form the largest group of the TGF (transforming growth factor)-β superfamily.52-55 They are so named for their osteo-inductive properties, and regulate differentiation of mesenchymal cells into components of bone, cartilage or adipose tissue.52 TGF-β/BMP ligand signal is mediated by heterodimeric receptor serine/threonine protein kinases (receptor types 1 and 2) and a family of receptor substrates (the Smad proteins) that move into the nucleus. Once activated, receptor kinases phosphorylate R-Smads (receptor-regulated Smads), and phosphorylated R-Smads then complex with C-Smads (common-mediator Smads). The complexes, which act as transcriptional regulators, then translocate into the nucleus.54 55 All Smads recognise and have low affinity interaction with the same sequence; high affinity interaction is facilitated by other transcriptional cofactors including OAZ, Mixer, and Milk.55 BMP-2 and BMP-7 induce the critical transcription factors Runx2 and Osterix in mesenchymal stem cells and promote osteoblast differentiation.56 57
RUNX2 and osterix
During development of the skeleton and formation of bone tissue, Wnt/β-catenin and TGF-β/BMP pathways modulate key transcriptional factor RUNX2(CBFA1/AML3) to induce the osteogenic phenotype.15-21 RUNX2, a member of the runt homology domain transcription factor family is essential for osteoblast differentiation and is also required for embryonic bone formation. The Runx family has three members, Runx1/Cbfa2/Pebp2aB, Runx2/Cbfa1/Pebp2aA and Runx3/Cbfa3/Pebp2aC. They all have a runt domain that is a DNA-binding homologue of Drosophila pair-rule gene runt.58 Runx proteins interact with transcriptional co-activator core binding factor b (Cbfb)/polyoma enhancer binding protein 2b (Pebp2b) forming a heterodimer; and Cbfb is also required for Runx2-dependent osteoblast and chondrocyte differentiation.19 20 59 Runx2-/- transgenic mice have complete lack of both endochondral and intramembranous ossification due to the absence of osteoblast differentiation.16 17 While Runx2 is essential for differentiation of mesenchymal cells into osteoblasts, both Runx2 and Runx3 are essential for chondrocyte maturation, and hence endochondral ossification.60 Runx2 however inhibits differentiation of mesenchymal cells into adipocytes and chondrocytes.60 61 Runx2 triggers the expression of major bone matrix protein genes, including the Col1a1, osteopontin and osteocalcin genes at an early stage of osteoblast differentiation and its actions sustain a supply of preosteoblasts.
Osterix, a member of the SP family of transcription factors is a second transcription factor that is essential for osteoblast differentiation.62 Osterix-/- and Runx2-/- mice have similar phenotypes; both show complete lack of endochondral and intramembranous ossification due to the absence of osteoblast differentiation. Osterix is believed to act downstream of Runx2, since mesenchymal cells from osterix-/- mice express Runx2, but osterix is not expressed in Runx2-/- mice.62 Osterix-/- mesenchymal cells retain the ability to differentiate into chondrocytes.62 Both RUNX2 and osterix expression in mature osteoblasts are modulated by intracellular immunophilin mediated calcineurin/NFATc1 pathway and enhance alkaline phosphatase and Col1a1 promoter activity.63
To summarise, β-catenin directs osteoblast differentiation at the preosteoblast stage, βcatenin/TCF1 enhances Runx2 expression and its promoter activity. Runx2 is required for mesenchymal progenitor cell differentiation into preosteoblasts, and for suppressing their differentiation into adipocytes and chondrocytes. Osterix and β-catenin act by directing the differentiation of preosteoblasts into immature osteoblasts and abolish the potential of preosteoblasts for differentiating into chondrocytes.
Runx2 transcriptional regulators
Many proteins interact with and modulate Runx2 activity. Cbfb is considered a critical regulator of Runx2, as it is required for DNA binding of Runx2. Nevertheless, Runx2 retains limited function even in the absence of Cbfb, implying that as yet unidentified factors may substitute for Cbfb. Runx2 activity is enhanced by a number of transcription factors, including ATF4, C/EBPb, C/EBPd, ETS1, Grg5, Menin, p204, Rb, Smad1, Smad5 and TAZ. It is inhibited by others including C/EBPd, Dlx3, Msx2, PPARg, Smad3, Stat1, TLE, Twist and Yes.64-66 Twist-1, a member of the basic helix-loop-helix (bHLH) family of proteins, is expressed in embryological mesodermal and cranial neural crest cells during embryogenesis.67 Expression of Twist-1 has been associated with the inhibition of differentiation for multiple mesenchymal cell lineages, including bone.68-70 Twist-1 interacts directly with Runx2, and this causes gene inactivation in osteoblast precursors.70 Id1, an internal dominant negative form of HLH transcription factor l, induces Twist-1 degradation and alleviates the inhibitory effect of Twist-1.71 Inhibition of BMP signalling by Twist-1 is enhanced by E-protein, indicating that Twist-1 and Id1 regulate differentiation of mesenchymal cell lineages by controlling BMP signalling.72 73
Osteocytes
Osteoblasts are encased by the matrix that they themselves synthesise and become osteocytes. Osteocytes, found dispersed throughout the bone matrix, are terminally differentiated cells that have much lower bone forming activity than osteoblasts, but constitute over 90% of adult bone cells.74 Despite their relative inactivity compared with osteoblasts, osteocytes play a central role in the determination and maintenance of bone structure.75-79 The bone matrix isolates osteocytes from each other and instead osteocytes interact with other osteocytes and bone cells by an elaborate network of osteocytes (dendritic) processes, which run inside lacunar cannaliculi. These processes may have the potential to stimulate bone resorption.78 79 It has long been recognised that mechanical stress induced by weight-bearing exercise increases osteoblast activity. Indeed, the absence of mechanical stimulation resulting from prolonged immobilisation or microgravity causes severe bone loss.80 81 There is an emerging consensus that osteocytes act as mechanosensory cells and lacuno-canaliculae carry signalling molecules that are responsible for maintenance of bone structure and mass.82-84 This model has been proposed to explain how mechanical loading induces biochemical transmission that promotes bone formation and remodelling.
Osteocytogenesis
The process of osteocyte differentiation or osteocytogenesis, whereby osteoblasts become encased in mineralised bone matrix is not yet understood, but there have been some important observations. The most abundant matrix protein in the osteocyte environment is type-I collagen. Osteocyte phenotype and the formation of osteocyte processes has been shown to be dependent on cleavage of type-I collagen.77 85 Osteocytes from metalloproteinase MT1-MMP (-/-) mice have a significantly reduced number and length of dendritic processes; MT1-MMP is a membrane-anchored proteinase that can cleave collagens type I, II, and III, fibrin, fibronectin, and other matrix molecules.77 85 Changes in osteocytes dendrite number have been suggested to effect osteocyte function and viability, as well as the mechanical properties of bone. The fluid flow shear stress-related osteocyte dendrite lengthening has been shown to be due to protein E11. E11 protein, which is also known as Gp38, podoplanin, RTI40, and T1{alpha}, plays a role in the formation of dendritic processes; its expression is increased in response to fluid flow.86
A number of matrix proteins, such as dentin matrix protein 1 (Dmp1), matrix extracellular phosphoglycoprotein or osteocytes/osteoblast factor 45 (MEPE/OF45), and sclerostin increase in expression once the osteoblast begins to transform into an osteocyte.87 89 DMP1, an acidic phosphorylated extracellular matrix protein, is a member of a family of proteins called SIBLING (Small; Integrin-Binding LIgand, N-linked Glycoprotein).90-92 Other members include MEPE and bone sialoprotein (BSP), osteopontin (OPN), and dentin sialophosphoprotein (DSPP); all bind hydroxyapatite and possess RGD sequence. OPN and DMP1 bind CD44, which interacts with the ERM (ezrin, radixin, moesin) family of adapter proteins that link to actin in the cytoskeleton.93 94 That DMP1 may have an important role in osteocyte maturation is evident from its high level of expression in osteocytes; particularly in the dendritic processes of these cells, and increased expression in response to mechanical loading.88 Osteocytes from DMP1-null mice show abnormalities in osteocyte morphology and severe impairment in mineralisation. DMP1 -/- mice and human autosomal recessive hypophosphataemic rickets display similar phenotypes, with raised FGF23 producing renal phosphate-wasting associated rickets. In contrast, MEPE null mice have raised osteoblast activity, increased bone mass, and are resistant to aging-associated trabecular bone loss, suggesting that it has an inhibitory role in bone formation.88 90 91
Bone mechanoreceptor cells
The transformation of mechanical stress into biochemical signals occurs in osteocytes and osteoblasts and involves a variety of membrane proteins, including integrins, connexins and stretch-activated ion channels. Potential biochemical intracellular signal mediators include IP3 and intracellular calcium, cAMP, guanine regulatory proteins and MAPK.95-99 Integrins are transmembrane dimers made of {alpha} and β subunits; the β subunit has also been reported in osteocytes.100 Fluid flow up-regulates β1 expression and activates {alpha}vβ3 which co-localises with c-src.101 Integrin signalling activates MAPK via β1 integrin and FAK dependent tyrosine phosphorylation in endothelial cells. FAK brings about MAPK activation via interaction with c-src, Grb2 and the small GTPase Ras.102 103 FAK activates the {gamma}1 isoform of PL, leading to intracellular calcium mobilisation. This pathway may be important in osteocytes since fluid induced calcium mobilisation has been shown to require PLC mediated IP3 release. Mechanical stimulation increases expression of connexins, membrane-spanning proteins that form regulated channels; this allows the direct exchange of small molecules with adjacent cells, resulting in intercellular communication between cells. It has been shown that mechanical stimulation increases expression of connexins.104 Connexins can also form regulatory hemi-channels between the cell and its extracellular environment. Shear stress leads to the modulation of release of nitric oxide and eicosanoid from osteoblasts and osteocytes, which may be due to activation of nitric oxide synthase.105 106
Osteoclasts
The respective bone resorbing and forming actions of osteoclasts and osteoblasts are finely coupled, so that in a healthy adult bone mass remains remarkably stable. There are however many disease states, such as osteoporosis, Paget's disease of bone, bone cancer metastases and inflammatory arthritis, where this fine equilibrium is disrupted due to a net increase in osteoclast bone resorbing activity. In view of the critical role of osteoclasts in diverse pathology, there has been immense effort aimed at understanding the biology of this unique cell.
Osteoclastogenesis
Osteoclasts are end-differentiated multinucleated cells of monocyte/macrophage lineage that are formed to carry out the unique function of resorbing bone matrix. It had been apparent for decades that in vitro differentiation of monocytes to osteoclasts could only take place when co-cultured with osteoblasts or other stromal cells.107 Subsequent investigations have shown that osteoclastogenesis is critically dependent on two key cytokines, namely RANKL (receptor activator of nuclear factor-{kappa}B ligand), also known as TRANCE (TNF-related activation-inducing cytokine) and M-CSF (monocyte-colony stimulation factor). M-CSF is thought to be critical for the proliferation of the osteoclast progenitors, while RANKL directly controls the differentiation process by activating RANK (receptor activator of nuclear factor-{kappa}B).108-111 It is now emerging that RANK cooperates with other receptors: OSCAR (osteoclast-associated receptor) and a triggering receptor expressed on myeloid cells (TREM)-2) (fig 5).112-114 OSCAR is an immunoglobulin-like receptor and, like RANKL, is involved in osteoblast-osteoclast interaction. OSCAR acts via an adaptor molecule FcR{gamma}, which has an immunoreceptor tyrosine-based activation motif (ITAM) that is critical for the activation of calcium signalling.115 DAP12, another ITAM-harbouring adaptor, is also involved in the formation and function of osteoclasts. There is emerging evidence for cooperation between the signalling that occurs via RANK, OSCAR/PIR-A and TREM-2. Stimulation of RANK and of the immunoglobulin-like receptors cooperatively phosphorylates ITAM.116 The precise mechanism as to how RANK induces osteoclastogenesis in cooperation with ITAM signalling is not fully understood. It is partly explained by the observation that RANKL leads to an increased phosphorylation of ITAM and increases expression of immunoglobulin-like receptors such as OSCAR, thereby augmenting the ITAM signal. In the absence of a functional DAP12 protein, FcR{gamma} mediated co-stimulatory signals can restore weak osteoclast differentiation. The identity of ligands that bind to DAP12- and FcR{gamma}-associated receptors, leading to the phosphorylation of their ITAM motif, is not known. The phosphorylated tyrosine residues in ITAM interact with the SH2 domain of Syk family kinase, and activate phospholipase C{gamma} (PLC{gamma}), leading to transient increase in intracellular calcium and the induction of nuclear factor of activated T-cells-2 (NFAT2, also known as NFATc1).117 118
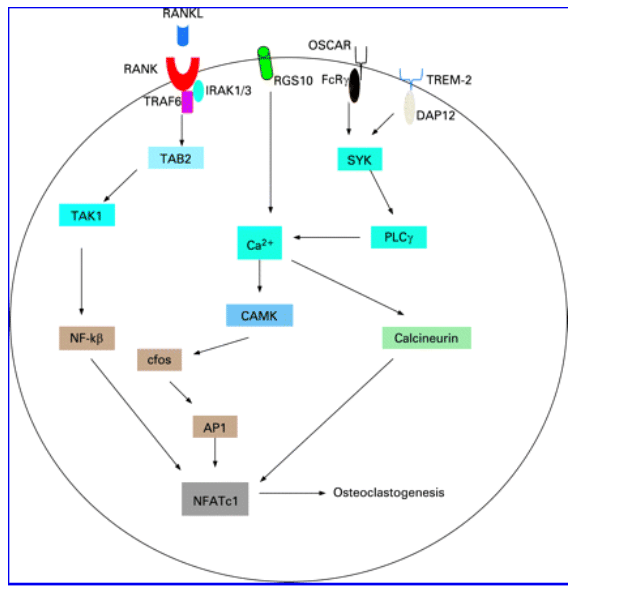
Figure 5 Molecular signalling involved in osteoclast differentiation. On activation RANK interacts with adapter protein TRAF6, which acts downstream to activate MAP kinases and NF-kβ. For the RANK signal to be effective, co-signalling from the immunoreceptors OSCAR and TREM2 is required. OSCAR and TREM interact with FcRa and with FcR{gamma} adapters that possess ITAM motifs and activate Syk kinases which in turn activate PLC{gamma}. The resulting mobilisation of Ca2+ from the intracellular stores activates calcineurin, which then dephosphorylates the transcription factor NFATc1. NFATc1 translocates to the nucleus and, acting in concert with c-fos, promotes expression of key osteoclast genes. The tyrosine kinase-family receptor c-fms and its ligand M-CSF are essential for the survival and proliferation of the progenitors cells.
RANK-RANKL interaction activates NFATc1 by an alternate pathway involving TRAF-6, c-Jun, c-Fos and p38.119-124 NFATc1 is critical for osteoclastogenesis as its over-expression abolishes the RANKL requirement for osteoclastogenesis, and NFATc1 (-/-) monocytes fail to form osteoclasts. RANK-RANKL signalling induces intracellular Ca2+ oscillations, which are regulated by a G-protein (RGS10). RGS10-null mice have impaired osteoclast differentiation resulting from the loss of [Ca2+]i oscillation regulation.125 This shows that RANKL-evoked G-protein (RGS10) mediated Ca2+ oscillations lead to calmodulin and calcineurin-dependent NFATc1 activation.125
Osteoclast bone resorption
Osteoclasts predominantly express {alpha}vβ3 integrin, which recognise the RGD-containing matrix proteins, such as OPN and BSP.126 The occupancy of aacgrvβ3 is associated with intracellular signalling, and increased intracellular Ca2+ and tyrosine phosphorylation and free radical production, resulting in osteoclast activation.127-129 Osteoclast-bone interaction leads to the formation of the osteoclast clear zone and the organisation of actin-based adhesion structures named podosomes. Podosomes are comprised of an F-actin core surrounded by vinculin, talin, actinin, fimbrin, gelsolin and vimentin.130 Extracellular matrix interaction with {alpha}vβ3 induces c-src dependent tyrosine phosphorylation, PYK2 activation and its translocation to podosomes. Some of the cytoskeletal changes are mediated by the Rho-GTPases.129 131-133 The process of osteoclast migration, polarisation and activation involves a number of other signalling molecules, such as Crkl,11, c-Cbl, p130 Cas, leupaxin PI3-K,12, RhoA,13 and PYK-2, and is also dependent on Wiskott-Aldrich syndrome protein (WASp).134 WASp has been identified in the sealing ring of resorbing osteoclasts, and co-localisation with c-Src, PYK2, PSTPIP and PTP-PEST. PTP-PEST coordinates the formation of the sealing ring and thus the bone-resorbing function of osteoclasts.134-136
A bone-resorbing osteoclast shows polarity; degradation products of collagen and other matrix proteins are transported by transcytosis to the basolateral membrane.137 138 However, the mechanisms involved in disposal of Ca2+ and phosphate, which accumulate within the resorption hemivacuole, are unclear. Ca2+ is continually transported to the basolateral surface and was postulated to be selectively disposed by channels and transporters.139-143
Osteoimmunology
Recent efforts are beginning to define the nature of the interactions between the immune and skeletal systems (fig 6). The realisation of the intricate nature of this interaction has been supported by the fact that a number of cell surface receptors, cytokines and signalling pathways serve a critical role both in the immune and skeletal systems. The close interaction between immune progenitors and skeleton is facilitated by their proximity in the bone marrow. Therefore, osteoimmunology, a discipline that encompasses the complex interactions and overlapping functions of the immune system and bone cells has emerged over recent years.
T cells
T cells rapidly up-regulate surface expression of RANKL on activation, and are therefore capable of regulating osteoclast differentiation and activity. Indeed, activated T cells have been shown to promote osteoclastogenesis in vitro, mediate bone erosion in inflammatory joint disease and play an essential role in the pathogenesis of osteoporosis in ovariectomised mice.144-146 In contrast, resting T cells actively suppress osteoclastogenesis in a cell:cell contact independent manner.147 RANKL is also expressed on memory T cells, but not on naive T cells, but the physiological significance of such differential expression is unclear.148 Apart from RANKL, T cells produce many other cytokines that modulate osteoclast activity and osteoclastogenesis; TNF-{alpha}, IL-6 and IL-17 are stimulatory, while IL-4, IL-13 and IL-10 are inhibitory.149 Since the range of cytokines produced by different T cell subsets show considerable variability, so does their effect on osteoclast number and activity. In addition to their effect on osteoclast function, T cells may also interact with osteoblasts directly and exert effects on bone turnover indirectly via their interactions with dendritic and B cells.150 151
B cells
The role of other immune cell types on bone metabolism is poorly understood, although emerging data suggest that the B cell may be important.151 For instance, it has been reported that B cell progenitors are capable of differentiating into osteoclasts in vitro, and that peripheral blood B cells suppress osteoclastogenesis. Recently, it has been shown that B cells are a source of osteoprotegerin (OPG), and that B-cell deficient mice have reduced bone mass, which can be reversed by adoptive transfer of B cells.151-153
B7-H3, a widely expressed member of the B7 family of the Ig superfamily of proteins, is expressed on the surface of the antigen-presenting cells; it down-regulates T cell functions by interacting with a yet unidentified membrane-bound ligand on T cells. B7-H3 and its unknown counter-receptor have been shown to play a positive regulatory role in bone formation as well as in bone-immune system interaction.154 155
Cytokines and growth factors
Many cytokines are important regulators of osteoclast and osteoblast function, and a majority of these cytokines are produced by cells of the immune system.156 The precise mechanisms of how these cytokines regulate osteoblast and osteoclast activities have not been fully defined; most act indirectly by regulating the interactions between osteoclasts and osteoblasts. For instance, many cytokines have been shown to regulate the expression of RANK and RANKL on osteoclast and osteoblasts respectively, or modulate the intracellular signalling mediated by RANK. In general, pro-inflammatory cytokines, such as IL-1, TNF-{alpha}, and IL-6, appear to act synergistically on osteoclastogenesis and promote osteoclast function.157-162 Table 1 summarises the effects of the various cytokines on bone metabolism and the possible underlying mechanism(s) of action.
TNF-{alpha} enhances RANKL-mediated stimulation of osteoclasts by activating NF-{kappa}B.158 TNF-{alpha} also enhances the mobilisation of osteoclastogenesis progenitors from bone marrow and inhibits osteoblastic activity.159 Like TNF-{alpha}, IL-1 activates NF-{kappa}B, which promotes osteoclast survival and induces RANKL expression on osteoblasts.160 There are two forms of IL-1, IL-1{alpha} and IL-1β, but their biological activities are indistinguishable; cells of monocytic lineage on activation predominantly produce IL-1β, while IL-1{alpha} is released by dying cells and is not normally found in the circulation.161 Osteoblasts and stromal cells are the main source of IL-6 in bone; the effect of IL-6 on bone resorption is mediated primarily via stimulation of osteoclasts.162 163 IL-17, a key cytokine that mediates autoimmune inflammatory arthritis, also augments osteoclastogenesis.164 In contrast, Th2 cytokines such as IL-4, IL-13 and IL-10 suppress osteoclastogenesis and osteoclast activation.165 Interestingly, a recent report suggests that IL-4 may play an important role in CD4+CD25+Foxp3+ regulatory T cell-mediated suppression of osteoclastogenesis.166 Another class of cytokines that inhibits osteoclastogenesis is the interferons. IFN-β suppresses RANKL-induced expression of c-Fos; IFNβ-/- mice have increased osteoclast activity and are osteoporotic.167
Osteoblast regulation of energy metabolism
Recent evidence shows that osteoblast function may contribute to obesity and glucose intolerance. This suggests that the skeleton exerts an endocrine regulation of energy metabolism and thereby may contribute to the onset and severity of metabolic disorders. Esp (also known as Ptprv), a receptor-like protein tyrosine phosphatase termed OST-PTP that is expressed in osteoblasts has been shown to regulate insulin secretion.168 Mice lacking Esp in osteoblasts have increased β-cell activity that protects them from induced obesity and diabetes. These phenotypes are corrected by deleting one allele of osteocalcin, suggesting that osteocalcin may also play a role. Osteocalcin null mice are glucose intolerant; osteocalcin promotes proliferation of pancreatic β-cells and adiponectin expression in adipocytes and β-cells.169
Osteoblast regulation of energy metabolism
Recent evidence shows that osteoblast function may contribute to obesity and glucose intolerance. This suggests that the skeleton exerts an endocrine regulation of energy metabolism and thereby may contribute to the onset and severity of metabolic disorders. Esp (also known as Ptprv), a receptor-like protein tyrosine phosphatase termed OST-PTP that is expressed in osteoblasts has been shown to regulate insulin secretion.168 Mice lacking Esp in osteoblasts have increased β-cell activity that protects them from induced obesity and diabetes. These phenotypes are corrected by deleting one allele of osteocalcin, suggesting that osteocalcin may also play a role. Osteocalcin null mice are glucose intolerant; osteocalcin promotes proliferation of pancreatic β-cells and adiponectin expression in adipocytes and β-cells.169
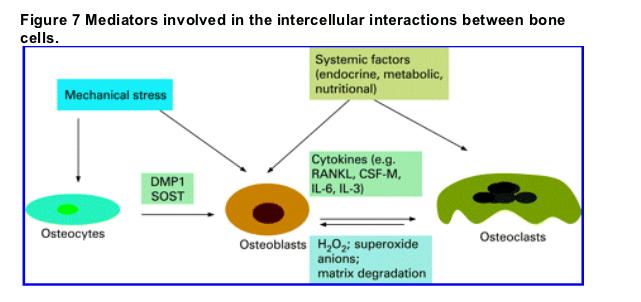
SUMMARY
Bone is a specialised tissue designed to provide sufficient strength to withstand mechanical forces, but yet to be light enough to allow mobility. It has a complex structure and undergoes constant remodelling. The remodelling process is undertaken by the basic multicellular unit of bone, which comprises osteocytes, osteoclasts and osteoblasts. They act in a coordinated manner to form or resorb bone as necessary. Their activation is controlled by mechanical forces, apoptosis, hormones, cytokines and local factors (fig 7). Osteocytes play a central role in the response to shear stress and micro-fracture, regulating bone remodelling via their dendritic processes and by apoptosis. The differentiation of progenitor cells into osteoblasts and osteoclasts is also under the control of complex regulatory mechanisms. There is growing evidence of interaction between the immune system and bone cells. A number of recent breakthroughs have shown that bones cells are not merely involved in bone remodelling and maintenance of bone mass, but have a number of critical functions. Among these newly described functions are the role of skeleton in energy metabolism, regulation of immunity and blood phosphate homoeostasis.
|
|
| |
| |
|
|
|