 |
|
|
| |
Low frequency K103N mutations are strongly associated with inadequate virologic responses to NNRTI-based therapy. sdNVP & K103N
|
| |
| |
Reported by Jules Levin
XVII HIV Drug resistance Workshop
June 10-14, 2008
Sitges, Spain
Gillian Hunt1, Ashraf Coovadia2, Elaine J. Abrams3, Gayle Sherman4, Tammy Meyers5, Johanna Ledwaba1, Cathal Seoighe6, Scott Hammer7, Lynn Morris1, Louise Kuhn8
1AIDS Research Unit, National Institute for Communicable Diseases, Johannesburg, South Africa 2Coronation Hospital, Wits Pediatric Clinics, University of the Witwatersrand, Johannesburg, South Africa
3International Center for AIDS Care and Treatment Programs, Mailman School of Public Health, Columbia University, New York, NY 4Department of Haematology, University of the Witwatersrand, Johannesburg, South Africa 5Harriet Chezi Clinic, Baragwanath Hospital, Wits Pediatric Clinics, University of the Witwatersrand, Johannesburg, South Africa
6Computational Biology Group, Institute of Infectious Disease and Molecular Medicine, University of Cape Town, Cape Town, South Africa 7Division of Infectious Diseases, College of Physicians and Surgeons, Columbia University, New York, NY
8Gertrude H. Sergievsky Center, and Department of Epidemiology, Mailman School of Public Health, Columbia University, New York, NY
Neverest1 was funded by BMS "Secure the Future" and the National Institute for Child Health and Human Development (NICHD)
AUTHOR CONCLUSIONS
Exposure to sdNVP >18 months earlier was not associated with reduced likelihood of achieving viral suppression by 24 weeks or with sustainability of suppression to 78 weeks following NNRTI-based therapy. Response to therapy was excellent in the cohort with >95% suppressing and 80% maintaining suppression to 78 weeks.
About 10% of sdNVP-exposed women had K103N mutations detected in RNA by AS-PCR only, whereas only 3% had K103N mutations detected in viral DNA.
K103N mutations detected by genotyping or AS-PCR are strongly associated with a compromised virologic response to NNRTI-based therapy. Most of these women initially suppressed; however this suppression was not sustained indicating a more than 3-fold increased risk of viral rebound by 78 weeks for those with detectable K103N pre-treatment compared to those without mutations. In other words, the consequences of underlying resistance mutations were not apparent within the first 6 months of therapy but clearly affected the durability of viral suppression after 6 months.
The relationship between pre-treatment mutations and inadequate viral response was also not absolute (~40% of those with mutations had durable responses) and about a third of those who we defined as having "inadequate viral responses" re-suppressed without a change in regimen following intensified adherence support.
sdNVP-exposed and unexposed women showed different patterns of mutations at virological failure especially M184V
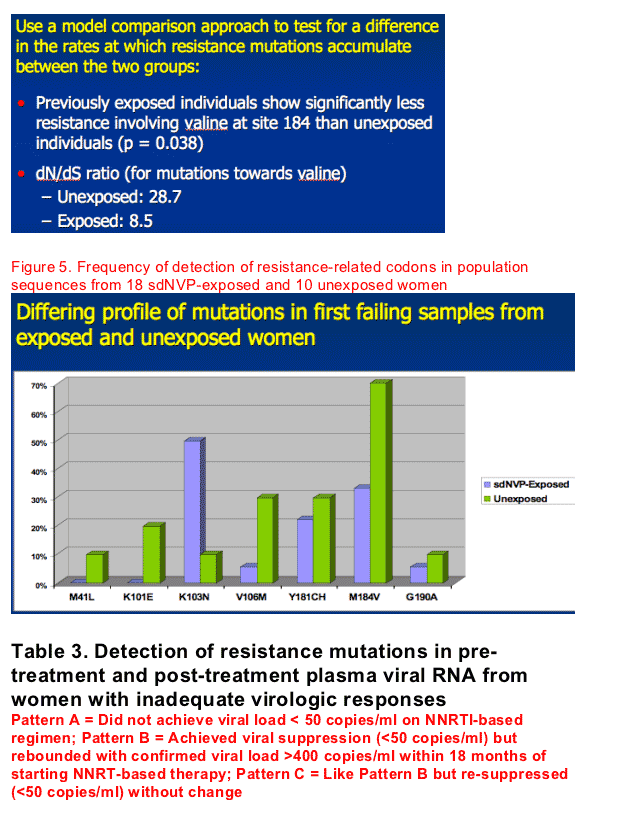
from Results below: Frequency comparisons of population sequences of the earliest failing sample revealed a different profile of mutations between the sdNVP-exposed and unexposed women (excluding the 2 sequences positive for K103N in the unexposed group by ASPCR and genotype): K103N was more common among the exposed (9/18, 50%) than the unexposed group (1/10, 10%), whereas M184V/I was more common among the unexposed (7/10, 70%) than the exposed group (6/18, 33%) (Figure 5 and Table 3).
BACKGROUND
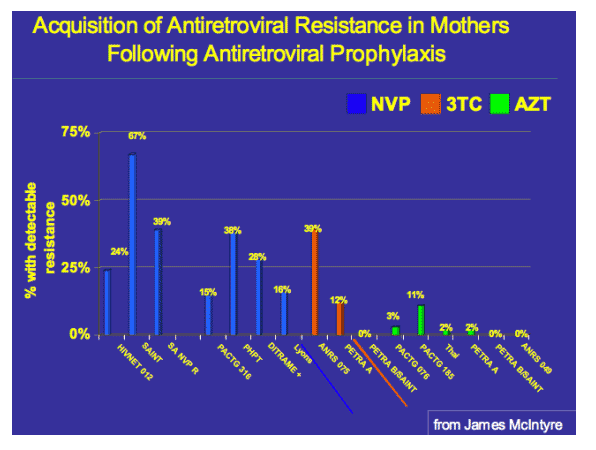
Single-dose Nevirapine (sdNVP) remains an inexpensive yet effective agent in significantly reducing mother-to-child HIV transmission in low resource settings, but is associated with the selection of viral resistance mutations, notably the K103N mutation of reverse transcriptase. Recently, two studies1,2 have shown that women initiated on a NNRTI-containing regimen after exposure to sd-NVP were less likely to achieve viral suppression compared to those who had not received sd-NVP as part of their PMTCT regimen; however this effect was not evident in women who initiated therapy after allowing a longer period of time to elapse. This difference is probably due to a rapid decline in NNRTI-associated mutations over time, as detected using population sequencing methodologies. Recently developed methods such as the allele-specific real-time PCR (AS-PCR) are capable of detecting lower frequencies of mutant variants than conventional genotyping assays, and reveal a persistence of these mutations at low levels in a small percentage of women following NVP-exposure. The consequence of these persisting minority populations of drug resistance mutations in virologic response to therapy has not been fully described.
METHODS
An observational epidemiologic study was conducted in Johannesburg, South Africa, in which initial and sustained virologic response to first-line therapy (d4T+3TC+NVP/EFV) was compared between 94 HIV-infected women previously exposed to sdNVP 18-36 months earlier and 60 previously pregnant women who were unexposed (Figure 1). All pre-treatment plasma and cell pellet samples were tested for the K103N mutation using AS-PCR and positive samples genotyped. Population sequencing and AS-PCR was performed on plasma samples from patients with inadequate virologic responses.

STATISTICAL ANALYSIS
Kaplan-Meier life tables were used to describe virologic outcomes and compared across groups with log-rank tests.
Standard codon models of evolution were adapted to a model-based approach for the detection of directional selection to a specific set of amino acids. The model was designed for sets of paired sequences, such as before and after exposure to antiretroviral therapy, and investigates the relationship between diversifying and directional selection and the evolution of drug resistance, and to identify novel drug resistance mutations3.
RESULTS
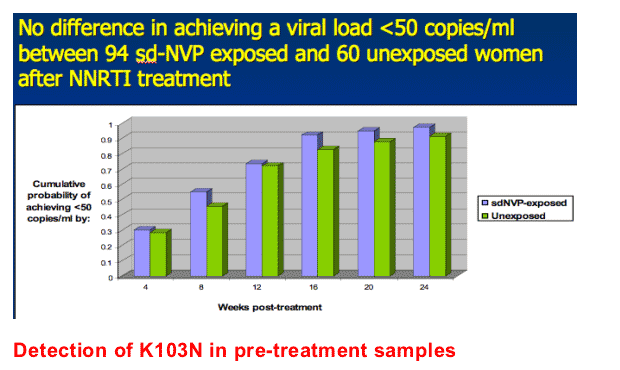
Almost all sdNVP-exposed and unexposed women achieved a plasma viral load <50 copies/ml by 24 weeks. The cumulative probability of achieving viral suppression by 24 weeks was 0.975 among sdNVP-exposed women and 0.913 among unexposed women (p=0.21) (Figure 2). Among women who achieved viral suppression with NNRTI-based therapy, there was no difference between sdNVP-exposed and unexposed women in the risk of viral rebound by 78 weeks (p=0.57).
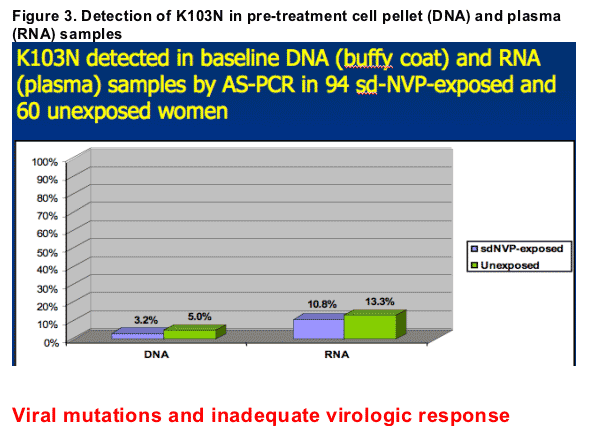
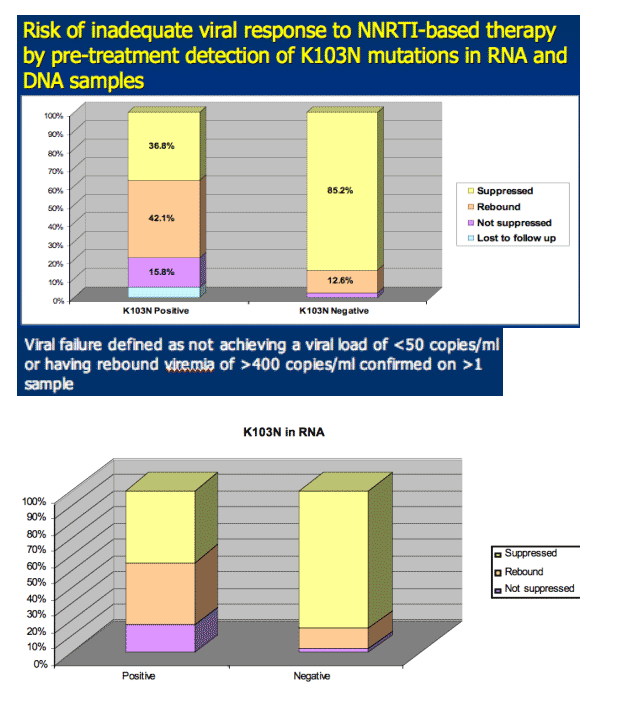
K103N mutations were detected by AS-PCR in 10/94 (10.6%) sdNVP-exposed women. Specifically, K103N was detected in 10/93 (10.8%) pre-treatment plasma (RNA) samples and in 3/94 (3.2%) cell pellet (DNA) samples.
K103N was also detected in 9/60 (15.0%) unexposed women: in 8/60 (13.3%) plasma (RNA) samples and in 3/60 (5.0%) cell pellet (DNA) samples (Figure 3).
Detection of K103N mutations by AS-PCR in pre-treatment samples was a strong predictor of virologic failure through 78 weeks of NNRTI-based therapy (defined as not achieving a viral load <50 copies/ml or having rebound viremia of >400 copies/ml confirmed on more than one sample).
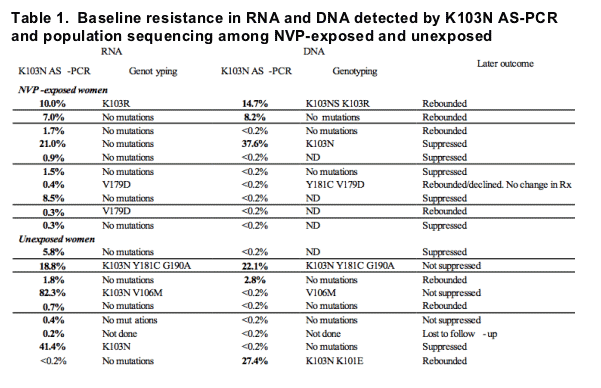
Eleven of 19 (57.8%) women with detectable K103N mutations in either viral DNA or RNA had poor virologic outcome - 3 did not suppress and 8 initially suppressed but rebounded (Figure 4 and Table 1).
However, the relationship between pre-treatment K103N and poor treatment outcome was not absolute: despite detectable K103N in pre-treatment samples: 7 (36.8%) women attained and sustained suppression through 78 weeks (1 was lost to follow-up).
The estimate by Kaplan-Meier methods of cumulative probability of inadequate virologic response by 78 weeks was 0.609 among women with pre-treatment K103N mutations vs. 0.151 among women without (p<0.001).
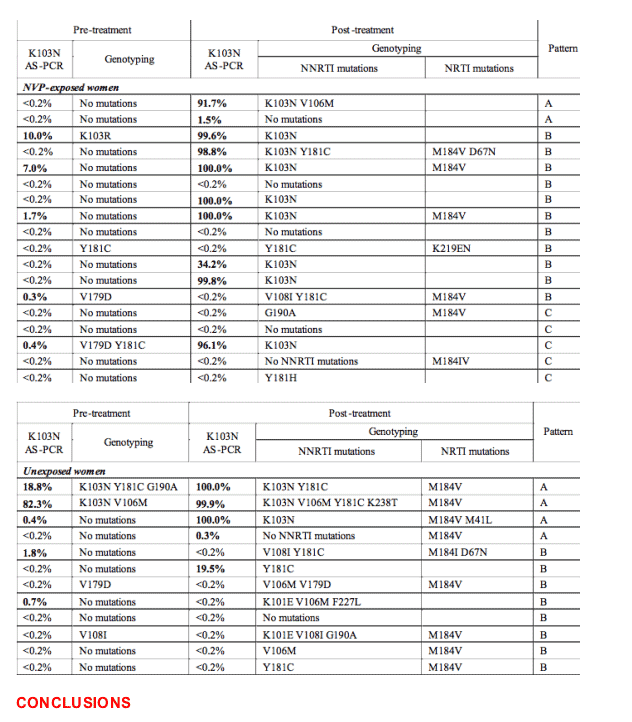
Population sequence data and clinical outcomes from the 10 sdNVP-exposed and 9 previously unexposed women positive for K103N by AS-PCR in pre-treatment samples are listed in Table 1.
However, out of the 30 women in both groups with inadequate virologic response, only 36.7% had K103N detected in pretreatment samples indicating that other minority populations need to be analysed.
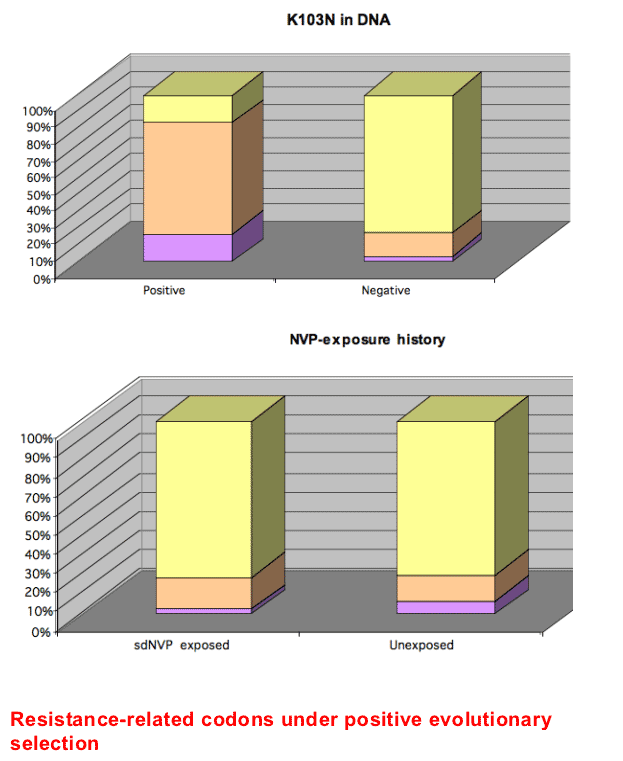
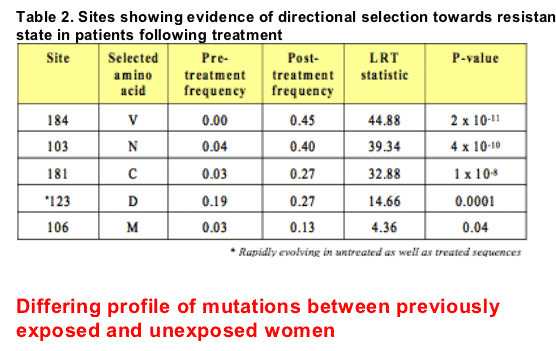
We applied a model-based approach of codon evolution to 30 pairs of sequences (i.e. pretreatment and earliest failing sample) from both groups of women to detect specific amino acid sites under directional selection. Mutations that were statistically shown to accumulate more rapidly than expected by chance, or in other words, are sites statistically shown to be under directional selection following exposure to NVP are listed in Table 2.
Frequency comparisons of population sequences of the earliest failing sample revealed a different profile of mutations between the sdNVP-exposed and unexposed women (excluding the 2 sequences positive for K103N in the unexposed group by ASPCR and genotype): K103N was more common among the exposed (9/18, 50%) than the unexposed group (1/10, 10%), whereas M184V/I was more common among the unexposed (7/10, 70%) than the exposed group (6/18, 33%) (Figure 5 and Table 3). The frequency of M184V in failing sequences was shown to be marginally significant between the two groups (p=0.06).
We further adapted the above model comparison approach to test for a difference in the rates at which certain resistance mutations accumulate between the two groups and showed that sdNVP-exposed individuals showed significantly less resistance involving valine at position 184 in RT than unexposed individuals: the dN/dS ratio for mutations towards valine was 28.7 for unexposed women and 8.5 for exposed women (p=0.038). The rate of accumulation of K103N was higher in the exposed women, although this was not statistically significant.
However, in both groups the majority had at least one NNRTI resistance mutation; 13/18 (72%) among sdNVP-exposed and 8/10 (80%) among unexposed women in their failing sample.
Exposure to sdNVP >18 months earlier was not associated with reduced likelihood of achieving viral suppression by 24 weeks or with sustainability of suppression to 78 weeks following NNRTI-based therapy. Response to therapy was excellent in the cohort with >95% suppressing and 80% maintaining suppression to 78 weeks.
About 10% of sdNVP-exposed women had K103N mutations detected in RNA by AS-PCR only, whereas only 3% had K103N mutations detected in viral DNA.
K103N mutations detected by genotyping or AS-PCR are strongly associated with a compromised virologic response to NNRTI-based therapy. Most of these women initially suppressed; however this suppression was not sustained indicating a more than 3-fold increased risk of viral rebound by 78 weeks for those with detectable K103N pre-treatment compared to those without mutations. In other words, the consequences of underlying resistance mutations were not apparent within the first 6 months of therapy but clearly affected the durability of viral suppression after 6 months.
The relationship between pre-treatment mutations and inadequate viral response was also not absolute (~40% of those with mutations had durable responses) and about a third of those who we defined as having "inadequate viral responses" re-suppressed without a change in regimen following intensified adherence support.
sdNVP-exposed and unexposed women showed different patterns of mutations at virological failure especially M184V
REFERENCES
1.Jourdain G. et al 2004. N Engl J Med 351:229-40.
2.Lockman S. et al, 2007. N Engl J Med 356:135-47
3.Seoighe C. et al, 2007. Mol Biol Evol 24(4):1025-31
|
| |
|
|
|
|
|