 |
|
|
| |
Transitions from CCR5 to CXCR4 use in the absence of antiretroviral drug pressure proceed incrementally and may occur through a multitude of genetic pathways: Monogram Report, Vs loop changes vary
|
| |
| |
Reported by Jules Levin
XVIII International HIV Drug
Resistance Workshop
Ft. Myers, Florida
June 9-13, 2009
W Huang1, E Coakley1, J Toma1, A Frantzell1, E Stawiski1, H
Schuitemaker2, CJ Petropoulos1 and AB van't Wout2
1Monogram Biosciences, South San Francisco USA.
2University of Amsterdam, Amsterdam,
The Netherlands
BACKGROUND:
The clinical relevance of the emergence of HIV subpopulations that
utilize the CXCR4 co-receptor is becoming increasingly important.
CXCR4 use is prognostic for disease progression, negatively predictive for
response to CCR5 antagonists, and represents a prevalent escape pathway
for CCR5-inhibitor treatment.
To advance our understanding of the acquisition of CXCR4 use, we characterized co-receptor utilization along with envelope clone sequences
derived from patient virus populations that evolved the ability to use CXCR4 in
the absence of antiretroviral drug pressure.
METHODS:
Longitudinal plasma samples were obtained from eight, treatment-naïve
subjects with previously documented tropism switches (NSI to SI) in the Amsterdam Cohort.
Envelope clones were isolated at different time points for each subject.
Co-receptor tropism was assigned by Trofile.
Envelope sequences were determined using conventional methods.
RESULTS:
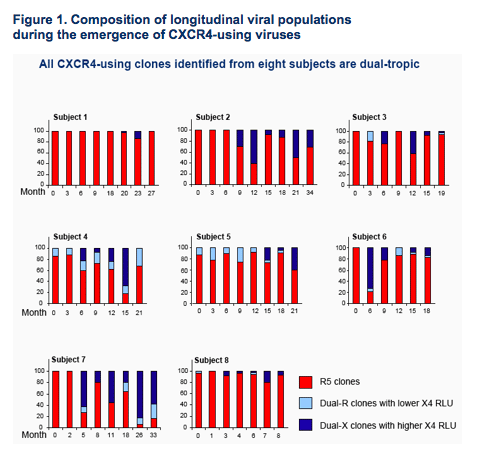
Virus populations at the time of the coreceptor switch were comprised of mixtures of R5- and dual-tropic clones (Fig.1)
The proportion of dual-tropic clones in virus populations and their ability to use
CCR5 and CXCR4 varied by subject (Fig.1). Dual-X clones that use CXCR4
efficiently were identified in all eight subjects; dual-R clones that use CXCR4
inefficiently appeared in six patients and presented either prior to, or concurrently with, dual-X tropic clones (Fig.1)
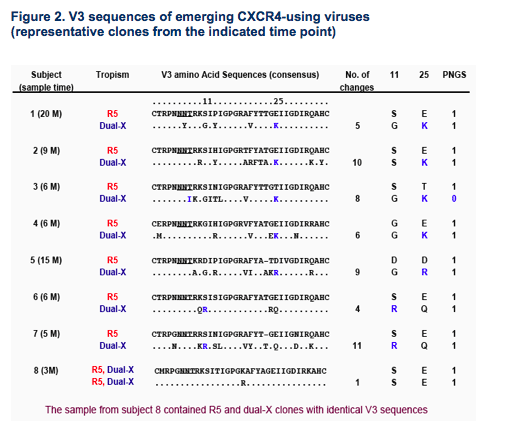
Generally, efficient CXCR4 use required multiple amino acid changes in the V3
loop, but few discernable patterns were shared among subjects (Fig.2).
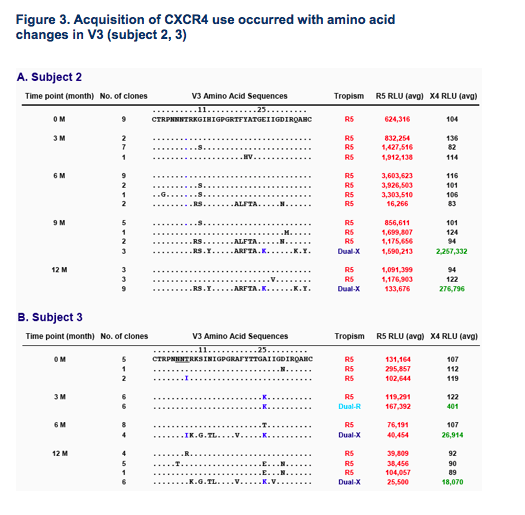
The presence of positively charged amino acids at positions 11/25 and the absence of potential N-linked glycosylation sites in V3 were associated with, but not unique to, CXCR4 use (Fig.2 and 3B).
V3 changes conferring CXCR4 use differed by subject both in composition and order of appearance (Fig 3A and 3B).
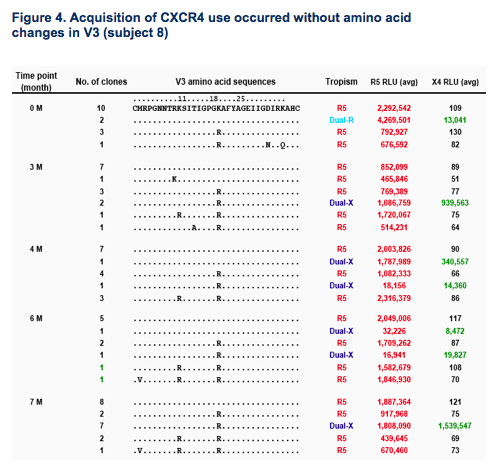
Efficient CXCR4 use can emerge in the absence of V3 sequence changes (Fig.4).
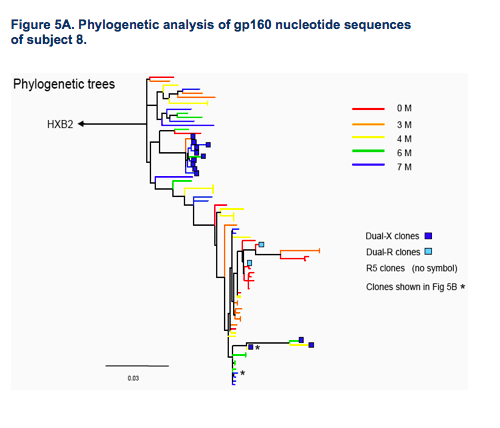
Emerging CXCR4-using variants can exhibit close genetic distance to R5 variants (Fig 5A).
Changes outside of the V3 region can confer efficient CXCR4 use (Fig.5B).
CONCLUSIONS:
Detailed characterization of emerging CXCR4-using variants in the virus
populations of eight individuals indicates that HIV uses a multitude of mutational pathways to transition from CCR5 to CXCR4 use.
Our observations indicate that CCR5 to CXCR4 transitions are incremental, not
quantum, and are consistent with the "R5, dual-R, dual-X" model to describe the
evolution of CXCR4 use from R5-tropic virus populations.

|
| |
|
|
|
|
|