 |
|
|
| |
Effect of Raltegravir-containing Intensification on HIV burden and T cell Activation in the Gut of HIV+ Adults on Suppressive Antiretroviral Therapy (ART) - reduced activation
|
| |
| |
Reported by Jules Levin
17th CROI, Feb 16-18 2010 SF
Steven Yukl1, Amandeep Shergill1, Sara Gianella2, Alex L. M. Choi1, Valerie Girling3, Michele Downing3, Harry Lampiris1, Huldrych F. Guenthard2, Joseph K. Wong1, Diane V. Havlir3, and the PLUS study group1. San Francisco VA Medical Center (SFVAMC) and University of California, San Francisco (UCSF), San Francisco, CA, USA2. University Hospital Zurich, Switzerland3. San Francisco General Hospital and University of California, San Francisco (UCSF), San Francisco, CA, USA
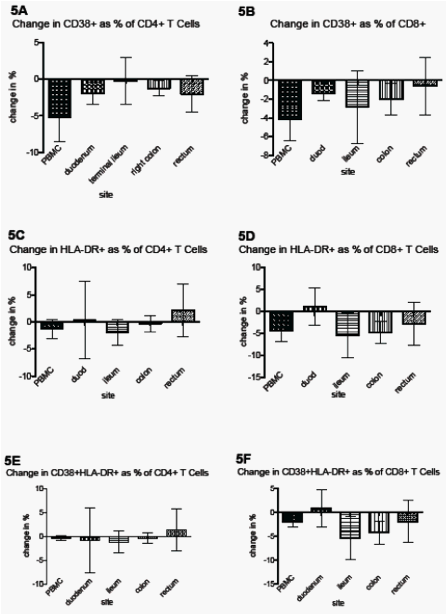
"T Cell Activation (Figure 5): Six of 7 participants had a decrease in CD38+ T cells as % of CD8+ T cells (5B) in PBMC (from mean of 40.61% to 36.43%; median change -4.84%) and colon (from mean of 89.90% to 87.90%; median -1.11%). Five and 6 (respectively) out of 7 also had a decrease in CD38+HLA-DR+ cells as % of CD8+ T cells (5D) in the PBMC (from 15.24 to 13.21; median -1.9%) and colon (from 46.94% to 42.63%; median -4.27%)."
Discussion
We evaluated the effect of raltegravir-containing ART intensification on viral burden, T cell reconstitution, and immune activation in the blood and 4 gut sites.
ART intensification did not reduce HIV RNA in the plasma, even though we intensified with one to two new agents and applied an assay detecting <0.5 copy of HIV per ml. This finding argues against ongoing viral replication in circulating lymphocytes as a major source of persistent plasma virus.
Intensification resulted in no consistent change in cell-associated HIV RNA in either blood (PBMC or CD4+ T cells) or in the duodenum, colon, or rectum. This finding suggests that most of the HIV RNA in the blood and these gut sites is not the result of ongoing replication that can be affected by short-term intensification with RLG-containing regimens.
In the ileum, which has the highest baseline HIV RNA/DNA ratio, HIV RNA declined in 5 of our study participants in conjunction with ART intensification. Supporting the interpretation that viral production was affected in these subjects, we also observed a decrease in CD8 immune activation markers and an increase in proportion of CD4+ T cells in the ileum.
ART intensification resulted in no consistent decrease in HIV DNA in the gut. Overall, the lack of change in gut HIV DNA with intensification suggests that most of this DNA represents latent or replication-incompetent provirus rather than the result of new infectious events that can be prevented by raltegravir.
In 2 subjects, an observed decrease in HIV DNA was reversed at week 16 in the CD4+ T cells but not the PBMC, suggesting that HIV DNA may have a different nature, origin, and/or stability in different cell types within the blood.
Study limitations include the small number of subjects, short period of intensification, possible confounding effects of the biopsies themselves (leakage of pro-inflammatory microbial substances), and the inherent difficulty in sampling infectious events that could occur in minute foci separated in both space and time (which reduces the value of negative results).
Summary
1. Our data suggest that multiple mechanisms are responsible for the persistence of HIV in patients on ART.
2. The lack of significant changes in viral RNA in blood and colon, duodenum and rectum implies a process other than ongoing replication, such as reactivation from latency or chronic persistence of virions and/or chronically-infected, transcriptionally-active cells.
3. At the same time, the changes in ileal HIV RNA, immune activation and CD4 counts in some patients are consistent with ongoing replication in these patients.
4. Future studies are needed to confirm the presence and site of ongoing replication and to identify other clinical factors that may predict which patients have ongoing replication.
5. While these patients may potentially derive benefit from intensification of ART, other strategies will be necessary for viral eradication.
Results
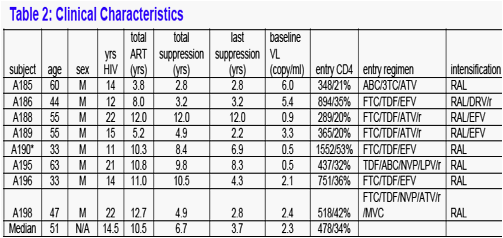
Study Population and Clinical Outcome (Table 2)
Seven participants (Table 2) completed the study; one participant (A190) withdrew for non-medical reasons before starting the study intervention.
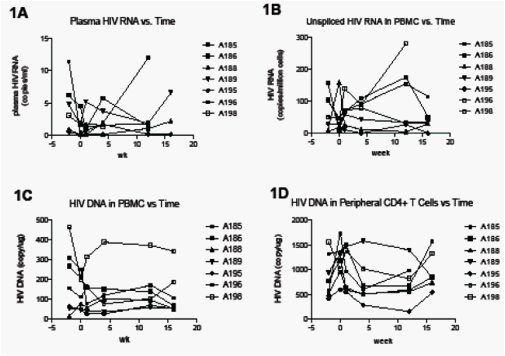
Plasma HIV RNA (Figure 1A)
Plasma HIV RNA was undetectable in all study participants using the standard Abbott assay, but was detectable from at least one of the two baseline time points in all participants using our high volume HIV RNA assay (median 2.4 copies/ml; range 0.5to 6 copy/ml). Intensification resulted in no consistent decrease in the plasma HIV RNA (Figure 1A).
Cell-associated HIV RNA in peripheral blood (Figure 1B)
ART intensification resulted in no consistent change in the Us HIV RNA in PBMC (Figure 1B), peripheral CD4+ T cells (data not shown), or non-CD4+ T cells.
Cell-associated HIV DNA in peripheral blood (Figure 1C-D)
Two participants, A185 and A186, appeared to have a decrease in HIV DNA in both PBMC (Figure 1C) and peripheral CD4+ T cells (1D) during the period of intensification. In both participants, the decrease in HIV DNA in PBMC was sustained after cessation of intensification (wk 16), whereas the decrease in HIV DNA in CD4+ T cells was almost completely reversed by week 16.
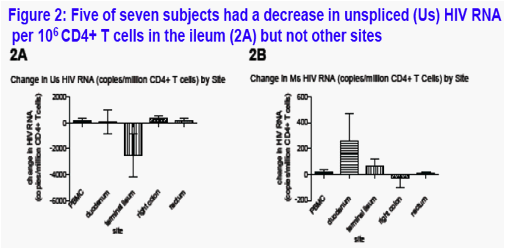
Cell-associated HIV RNA in gut (Figure 2A-B)
There was no significant change in Us HIV RNA/106CD4+ T cells in blood or the duodenum, right colon and rectum (Figure 2A). In the ileum, 5 of 7 participants had a decrease in Us RNA/106CD4+ T cells (2A) from mean of 3438 to 682 copies/106CD4+ T cells (median change: -576 copies/106CD4+ T cells). There was no appreciable change in Ms RNA normalized to total cells or to CD4+ T-cells (Figure 2B).
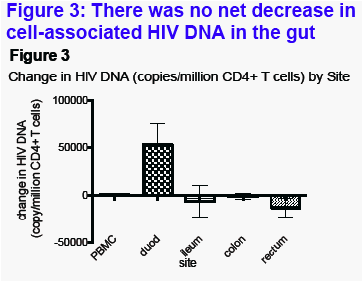
Cell-associated HIV DNA in the gut (Figure 3)All seven participants had an increase in HIV DNA/106 CD4+ T cells in the duodenum (Figure 3). There was no consistent change in the other gut sites or the blood.
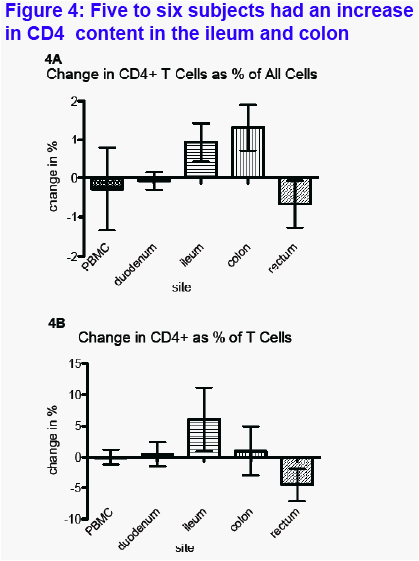
CD4+ T Lymphocytes (Figure 4 A-B)Six of 7 participants had an increase in CD4+ T cells in the terminal ileum (from a mean of 1.93% to 2.85%; median change +0.49% [Figure 4A]) and in the right colon (from mean of 2.55% to 3.85%; median change +1.13%). Five of 7 participants had an increase in CD4% in the ileum (from mean of 27.46% to 33.52%; median change +9.88%).
T Cell Activation (Figure 5)
Six of 7 participants had a decrease in CD38+ T cells as % of CD8+ T cells (5B) in PBMC (from mean of 40.61% to 36.43%; median change -4.84%) and colon (from mean of 89.90% to 87.90%; median -1.11%). Five and 6 (respectively) out of 7 also had a decrease in CD38+HLA-DR+ cells as % of CD8+ T cells (5D) in the PBMC (from 15.24 to 13.21; median -1.9%) and colon (from 46.94% to 42.63%; median -4.27%).
ABSTRACT
Background: Studies of ART intensification in suppressed patients fail to show consistent reductions in plasma HIV RNA. We hypothesized that in patients receiving intensification, reductions in HIV RNA and in immune activation may be seen in the gut.
Methods: In 7 HIV+ men with viral load <40 copies/mL for 3 to 12 yrs and a CD4 count >200, we performed a pilot study of 12 week intensification with raltegravir (RLG) alone (n = 4), RLG + EFV (n = 2), or RLG + DRV/RTV (n = 1). Gut cells were obtained by upper and lower endoscopy with biopsies from 4 gut sites (duodenum, ileum, colon, and rectum) at time 0 and 12 wks. Study outcomes included plasma HIV RNA (modified Abbott assay), HIV DNA from peripheral blood mononuclear cells and 4 gut sites (real time PCR), HIV RNA from peripheral blood mononuclear cells and 4 gut sites (real time RT PCR), T cell subsets, and activation markers (flow cytometry). Pre- and post-intensification values were compared using the paired Wilcoxon signed rank test.
Results: HIV was detectable in plasma (RNA) and in peripheral blood mononuclear cells (DNA and unspliced RNA) in all patients at baseline (medians: 2.3 copies/mL, 591 copies/106 cells, 44.4 copies/106 cells). HIV DNA was detectable in the duodenum in 63% of patients (median 61 copies/106 cells) and in the ileum, colon, and rectum in 100% of patients at baseline (medians: 370, 473, and 545 copies/106 cells). Unspliced HIV RNA was detectable in each gut site in the majority (63 to 88%) of patients at baseline (median: 8, 24, 7, and 10 copies/106 cells in duodenum, ileum, colon, and rectum). Intensification resulted in no consistent change in HIV RNA in the plasma, peripheral blood mononuclear cells, or gut, though there was a trend (5 of 7) towards decreased unspliced HIV RNA per 106 CD4+ T cells in the ileum (from mean of 3438 to 682). There was a trend towards decreased activation (CD38+, HLA-DR+, and dual+) of CD4+ and CD8+ T cells in all sites, which was greatest for CD8+ T cells in the ileum (mean of 25.6 to 20.2% CD38+HLA-DR+) and in peripheral blood mononuclear cells. There was also a trend towards increased CD4 as % of all cells (6 of 7; from a mean of 1.9 to 2.9%) and as % of T cells (5 of 7; from 27.5 to 33.5%) in the ileum.
Conclusions: In suppressed patients, most HIV RNA in the plasma, peripheral blood mononuclear cells, and gut is not the result of ongoing replication that can be reduced by short-term intensification with raltegravir. However, intensification reduced HIV RNA, reduced immune activation, and increased CD4+ T cells in the ileum, suggesting that the ileum may support ongoing productive infection in some patients on ART, even if the contribution to plasma RNA is not discernible.
Introduction
Although ART can reduce plasma HIV RNA levels below 40 copies/ml (suppression), HIV persists as latent provirus, low-level plasma RNA, and cell-associated HIV RNA in peripheral blood mononuclear cells (PBMC) and lymphoid tissues. It is unclear whether the residual HIV RNA results solely from reactivation of latent infection or whether there is a contribution from cryptic, low-level but continuous rounds of new infection (ongoing replication) or chronically-productive cells. This question is critically important for strategies aimed at eradication.
There is evidence both for and against ongoing replication. One approach to assess for ongoing replication is by intensification of suppressive ART with one or more additional agents whose activity affects a distinct part of the viral life cycle. Several recent studies of ART intensification in patients on contemporary ART with plasma HIV<40 copies/ml have failed to show a reduction in plasma HIV RNA. However, the plasma may not reflect changes elsewhere in the body, especially the lymphoid tissues, which are the major site of HIV infection in untreated patients.
Given the role of the gut and the gut-mucosal lymphoid compartment as a reservoir for HIV, we hypothesized that intensification may result in reductions in HIV RNA and/or immune activation that could be detected in the gut. Because HIV levels and immune environments may vary throughout the gut, we sought to examine this question using samples from different regions of the gut.
We performed a pilot prospective study of ART intensification with raltegravir (RLG) alone or in combination with a protease inhibitor [PI] or non-nucleoside reverse transcriptase inhibitor [NNRTI].
Gut tissue and cells were obtained by upper and lower endoscopy with biopsies from the duodenum, terminal ileum, right colon, and rectum at baseline (week 0) and 12 weeks after intensification. Study outcomes included plasma HIV RNA, cell-associated HIV RNA (both unspliced and multiply spliced) fromPBMC and 4 gut sites, cell-associated HIV DNA from PBMC and 4 gut sites, T cell subsets, and activation markers.
Methods
Patient Recruitment
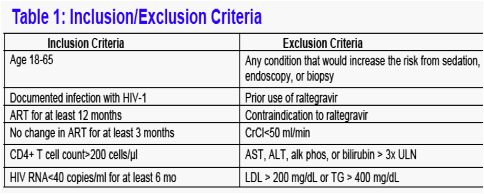
Inclusion and exclusions criteria are shown in Table 1. All patients who met these criteria were offered enrollment in the study. 8 elected to enter and 7 completed the study.
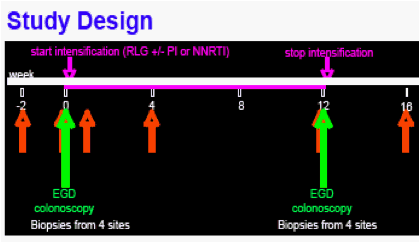
Study design
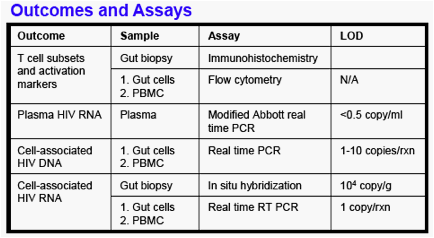
All participants had baseline upper and lower endoscopy with 7-10 biopsies each from the duodenum, terminal ileum, right colon, and rectum, followed by intensification (with raltegravir +/-study PI or NNRTI) for 12 weeks and repeat endoscopy with biopsies at week 12. HIV levels and immune markers were measured in intact biopsies and gut cells obtained by digestion with collagenase . Please see Study Design and Outcomes and Assays (above right).
Processing of Gut Biopsies
At least 6 biopsies from each site were separated into single cells by collagenase digestion and needle shearing. Aliquots were saved for flow cytometry and measurement of cell-associated HIV DNA or RNA by PCR. Intact biopsies were fixed overnight in 4% paraformaldehyde or Streck's cell preservative.
Processing of Blood
PBMC were isolated by centrifugation on Ficoll. 60 million PBMC were used to isolate CD4+ cells by negative selection using the Dynabeads Untouched Human CD4+ T Cell Kit (Invitrogen). Aliquots were saved for flow cytometry and measurement of cell-associated HIV DNA or RNA by PCR.
High Volume Plasma HIV RNA
We also measured plasma HIV RNA using a new assay that involves pelleting of plasma HIV virions from up to 30ml of plasma by high speed centrifugation on a density cushion. Plasma was diluted 1:1 with PBS, divided in two equal aliquots, layered onto 6% iodixanol, and spun at 20,000 rpm (47,810g) for 3hrs at 4C with deceleration 0. Viral pellets were resuspended in PBS, combined, and brought up to a final volume of 1ml. HIV RNA was then measured using the Abbott real time assay.
Cell-associated HIV DNA by Real Time PCR
Three replicates of 500ng of DNA from each sample were used in an in-house real time Taqman PCR assay for HIV DNA that uses primers and a Lock nucleotide acid (LNA) probe from the Gag region (detection limit=42% for 1 copy, 83% for 5 copies, and 95% for 10 copies). External standards (1e5 to 1e0) were prepared from DNA extracted from serial dilutions of known numbers of 8E5 cells. HIV DNA copy numbers were calculated from the Ct values and normalized by μg total DNA (1 μg DNA=160,000 cells) to get copies/106total cells. To account for wide variations in the number of CD4+ T cells in different samples, results were also normalized by the % of CD4+ T cells (measured by flow cytometry) and expressed as copies/106CD4+ T cells.
Cell-associated HIV RNA by Reverse Transcriptase Real Time PCR
Amplification primers and detection probes were custom designed for each study subject based on the sequence of the prevailing viral quasispecies (in peripheral CD4+ T cells) from that subject. Unspliced RNA (UsRNA) was measured using primers from the polregion (2536-2562 and 2634-2662). Multiply spliced tatRNA (MsRNA) was measured using primers from tatexon 1 (5956-5979) and tat/revexon 2 (8433-8459). Total multiply spliced RNA (TMsRNA) was measured using primers from revexon 1 (6012-6045) and tat/revexon 2 (8433-8459). Real time reverse transcriptase PCR was done under conditions described previously . Copy numbers were normalized to the cellular input as determined by UV spectrophotometry (assuming that 1ng RNA correspond to 1000 cells) and real time PCR for GAPDH. Results were further normalized to the % of CD4+ T cells (measured by flow cytometry).


|
| |
|
|
|
|
|