| |
NS5A HCV Drug Study Results, Genotypic and phenotypic analysis of variants resistant to hepatitis C virus nonstructural protein 5A replication complex inhibitor BMS-790052 in Humans: In Vitro and In Vivo Correlations
|
| |
| |
Download the PDF here
Hepatology December 2011
AASLD: Daclatasvir (DCV; BMS-790052), an NS5A Replication Complex Inhibitor, in Combination With Peginterferon Alfa-2b and Ribavirin in Japanese Treatment-Naïve and Nonresponder Patients With Chronic HCV Genotype 1 Infection - (11/10/11)
AASLD: Combination Therapy of Treatment-Naïve and Nonresponder Patients With HCV Genotype 1 Infection With Daclatasvir (DCV; BMS-790052), an NS5A Replication Complex Inhibitor, in Combination With Peginterferon Alfa-2a and Ribavirin - (11/10/11)
AASLD: Dual Oral Combination Therapy with the NS5A Inhibitor Daclatasvir(DCV; BMS-790052) and the NS3 Protease Inhibitor Asunaprevir(ASV; BMS-650032) Achieved 90% Sustained Virologic Response (SVR12) in Japanese HCV Genotype 1b-Infected Null Responders - (11/08/11)
ICAAC: BMS-790052 Plus Peginterferon Alfa and Ribavirin Demonstrated up to 83% Sustained Virologic Response 24 Weeks Post-Treatment (SVR24) in Phase II Study of Genotype 1 Hepatitis C Patients - (09/19/11)
Multiple ascending dose study of BMS-790052, an NS5A replication complex inhibitor, in patients infected with hepatitis C virus genotype 1 - (08/16/11)
First Report of SVR12 for a NS5A Replication Complex Inhibitor ...
www.natap.org/2011/EASL/EASL_13.htm
Apr 3, 2011 - BMS-790052 is a potentially first-in-class, highly selective HCV NS5A ... BMS-790052 inhibits HCV RNA replication through the NS5A protein
HCV Cured With out Peginterferon/ribavirin with 2 oral HCV drugs BMS790052+BMS650032: Quadruple Therapy With BMS-790052, BMS-650032 and Peg-IFN/RBV for 24 Weeks Results in 100% SVR12 in HCV Genotype 1 Null Responders: "HCV infection can be cured without interferon & ribavirin: 2 orals BMS790052+BMS650032" - (04/02/11)
Hepatology December 2011
Robert A. Fridell,1 Chunfu Wang,1 Jin-Hua Sun,1 Donald R. O'Boyle II,1 Peter Nower,1 Lourdes Valera,1
Dike Qiu,2 Susan Roberts,1 Xin Huang,3 Bernadette Kienzle,3 Marc Bifano,4 Richard E. Nettles,4 and Min Gao1
"Proof of concept for BMS-790052 has been achieved clinically, where its exceptional in vitro potency translated to an in vivo effect in a single-ascending dose study.....combination therapy will suppress the selection of resistant variants........In the multiple-ascending dose (MAD) study reported here, a total of 24 chronically infected patients (4 active patients per cohort) were treated with BMS-790052 at 1, 10, 30, 60, and 100 mg once-daily or 30 mg twice-daily for 14 days. The treated patients generally experienced rapid, marked viral load declines.....However, HCV RNA remained detectable in all genotype 1a-infected patients, and viral breakthrough was observed during the course of treatment in the majority of these patients. In contrast, viral breakthrough was observed less often in patients infected with HCV genotype 1b, and, in several patients, HCV RNA dropped below the level of quantitation (<25 IU/mL).6.....For genotype 1a, the major resistance substitutions observed in the study delivered greater resistance than single substitutions in genotype 1b.......Only a single-nucleotide change (UAU or UAC to AAU or AAC) at residue 93 (Tyr to Asn) of genotype 1a NS5A is required for HCV to acquire clinical resistance to BMS-790052 (Table 2). Furthermore, through either accumulation or novel mutation, linked substitutions emerged, such as Q30R-H58D (patient S, 100-mg cohort; Table 3E), which conferred a high level of resistance.....To maximize the anti-HCV response and minimize resistance, combination therapy, similar to current HIV treatment, could be used to enhance the resistance barrier. During combination therapy, variants with multiple substitutions, generally accompanied by reduced fitness, are required to generate clinical resistance. Our in vitro data show that BMS-790052-resistant variants remain fully sensitive to inhibitors of the NS3 protease and the NS5B polymerase,5 demonstrating the potential for effective combination treatment with DAAs that have different mechanisms of actions and targets."
Abstract
The NS5A replication complex inhibitor, BMS-790052, inhibits hepatitis C virus (HCV) replication with picomolar potency in preclinical assays. This potency translated in vivo to a substantial antiviral effect in a single-ascending dose study and a 14-day multiple-ascending dose (MAD) monotherapy study. However, HCV RNA remained detectable in genotype 1a-infected patients at the end of the MAD study. In contrast, viral breakthrough was observed less often in patients infected with genotype 1b, and, in several patients, HCV RNA declined and remained below the level of quantitation (<25 IU/mL) through the duration of treatment. Here, we report on the results of the genotypic and phenotypic analyses of resistant variants in 24 genotype 1-infected patients who received BMS-790052 (1, 10, 30, 60, and 100 mg, once-daily or 30 mg twice-daily) in the 14-day MAD study. Sequence analysis was performed on viral complementary DNA isolated from serum specimens collected at baseline and days 1 (4, 8, and 12 hours), 2, 4, 7, and 14 postdosing. Analyses of the sequence variants (1) established a correlation between resistant variants emerging in vivo with BMS-790052 treatment and those observed in the in vitro replicon system (major substitutions at residues 28, 30, 31, and 93 for genotype 1a and residues 31 and 93 for genotype 1b); (2) determined the prevalence of variants at baseline and the emergence of resistance at different times during dosing; and (3) revealed the resistance profile and replicative ability (i.e., fitness) of the variants. Conclusion: Although resistance emerged during monotherapy with BMS-790052, the substantial anti-HCV effect of this compound makes it an excellent candidate for effective combination therapy. (HEPATOLOGY 2011)
The hepatitis C virus (HCV) nonstructural protein 5A (NS5A) is a multifunctional protein with key roles in HCV replication. NS5A has also been implicated in the modulation of cellular signaling pathways.1, 2 Because it is required in vivo and in vitro for viral replication and has no known human homologs, NS5A provides an attractive target for therapeutic intervention.3
BMS-790052 is a potent HCV NS5A replication complex inhibitor, with 50% effective concentration (EC50) values of 9 and 50 pM against genotype 1b and 1a replicons, respectively.4, 5 It is also potent against live virus (genotype 2a, JFH-1), with an EC50 of ~28 pM.4 BMS-790052 has broad genotype coverage, with EC50 values ranging from pM to low nM for replicons with NS5A sequences derived from genotype 2a, 3a, 4a, and 5a.4
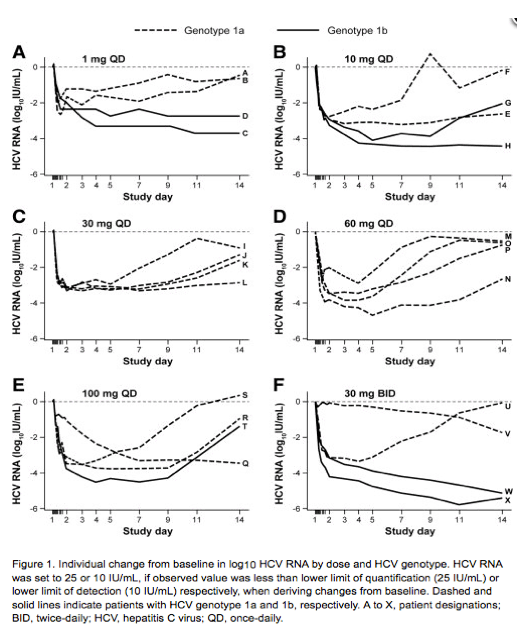
Proof of concept for BMS-790052 has been achieved clinically, where its exceptional in vitro potency translated to an in vivo effect in a single-ascending dose study.4 In this study, marked HCV RNA decline (~2.9 log10) was needed for detection of resistant variants, suggesting that the variants were likely present as preexisting minor quasi species and were uncovered by the efficient suppression of wild-type virus.4 However, the ability of BMS-790052 to further suppress viral replication with continuous dosing could not be assessed in the single-ascending dose study. In addition, analysis of specimens from the single-ascending dose study did not reveal whether the resistance detected clinically correlates with resistance observed in the in vitro replicon system. In the multiple-ascending dose (MAD) study reported here, a total of 24 chronically infected patients (4 active patients per cohort) were treated with BMS-790052 at 1, 10, 30, 60, and 100 mg once-daily or 30 mg twice-daily for 14 days. The treated patients generally experienced rapid, marked viral load declines. However, HCV RNA remained detectable in all genotype 1a-infected patients, and viral breakthrough was observed during the course of treatment in the majority of these patients. In contrast, viral breakthrough was observed less often in patients infected with HCV genotype 1b, and, in several patients, HCV RNA dropped below the level of quantitation (<25 IU/mL).6
The primary aims of the genotypic and phenotypic analysis of specimens from the MAD study were to (1) determine whether clinically relevant resistant variants identified during 14 days of monotherapy with BMS-790052 would correlate with resistance selected in vitro; (2) assess the prevalence of variants at baseline and the emergence of resistance at different times during dosing; and (3) examine the resistance profile and replicative ability (i.e., fitness) of variants. This article describes the viral variants identified by population sequencing and the phenotypic analysis of these variants using transient replicon replication assays.
Because there is a large population of viral quasi species preexisting in infected individuals, variants that confer resistance to antiviral agents can be rapidly enriched and selected during antiviral treatment. BMS-790052, like other direct-acting antiviral (DAA) agents,7-10 is, therefore, unlikely to be used as monotherapy. Variants with high levels of resistance to BMS-790052, which were identified in this MAD study, were evaluated for susceptibility to other DAA agents. Because no cross-resistance has been observed with other DAAs, our results suggest that BMS-790052 is an excellent candidate for combination therapy.
DAA, direct-acting antiviral; EC50, 50% effective concentration; HCV, hepatitis C virus; MAD, multiple-ascending dose; NS5A, nonstructural protein 5A.
Patients and Methods
Compounds.
The structure of the HCV NS5A inhibitor, BMS-790052, has been described previously.4, 5
Genotypic Analysis of Clinical Specimens.
Genotypic analysis of clinical specimens has been previously described.4 To determine the relative sensitivity of detecting sequence variants, reconstitution experiments were performed with DNA mixtures containing both wild-type and resistant (Y93H) variants. Mixtures of wild-type and Y93H DNA, at ratios of 100:0, 95:5, 90:10, 80:20, and 60:40, were sequenced. The experiment revealed that the variant could be readily detected at 20% of the wild-type population (results not shown).
In Vitro Analysis of Replicon Variants.
Substitutions were introduced into HCV reference replicons (genotype 1b, con1; genotype 1a, H77c), as previously described.4, 5 Inhibitor sensitivities and replicative ability (i.e., fitness) were assessed in transient replication assays, as previously described.4
Study Design and Clinical Specimens.
To examine safety, pharmacokinetics, and antiviral effect across the potential clinical dose range, BMS-790052 was dosed as monotherapy in a double-blind, placebo-controlled, sequential panel, MAD study in patients chronically infected with HCV genotype 1. Six dose regimens were evaluated (1 mg once-daily, 10 mg once-daily, 30 mg once- or twice-daily, 60 mg once-daily, and 100 mg once-daily). Five patients in each panel were randomized to receive a 14-day course of orally administered BMS-790052 or placebo in a ratio of 4:1; thus, a total of 24 patients received BMS-790052.6 The study was approved by the institutional review boards in all study centers and conducted in accord with good clinical practice and the ethical principles that have their origin in the Declaration of Helsinki.6 Informed written consent was obtained from all patients. HCV RNA levels were determined using the Roche Cobas TaqMan HCV Test, v2.0 (lower limit of quantification, 25 IU/mL; lower limit of detection, 10 IU/mL; Roche, Pleasanton, CA) at baseline and days 1 (2, 4, 6, 8, 12, 16, and 20 hours post-first dose), 2, 3, 4, 5, 7, 9, 11, 14, 15, 16, 17, 21, and 28. Viral breakthrough was defined as an HCV RNA increase by at least 0.5 log10 after HCV RNA nadir while receiving BMS-790052. Serum specimens were collected for potential genotypic analysis at baseline and days 1 (4, 8, and 12 hours post-first dose), 2, 4, 7, and 14. After amplification of the NS5A coding region, a genotypic analysis was performed by population sequencing to determine the emergence of viral variants after the administration of multiple doses of BMS-790052. The complete study design and resistance analysis methodology have been described elsewhere.5, 6
During the phenotypic analysis of NS5A variants, DAA agents, such as an NS3 protease inhibitor and/or an NS5B polymerase inhibitor, were used as controls for all analyses. Consistent with our previous report,5 NS5A variants are susceptible to these agents in the replicon system (Table 4), suggesting that these variants would likely be suppressed by combination therapy.5
Discussion
BMS-790052 is a novel HCV replication complex inhibitor, and the 14-day monotherapy study provided a unique opportunity not only to study the anti-HCV effect, but also to investigate the emergence of resistance. Studies of in vitro resistance to BMS-790052 using different levels of selective pressure and different replicon cell lines have been described.5 All amino acid substitutions associated with resistance to this class of inhibitors have mapped to the N-terminal region of NS5A.4, 5, 11 Patients receiving BMS-790052 monotherapy in the MAD study generally experienced rapid, marked viral load declines at early time points. However, viral breakthrough, especially in patients infected with genotype 1a virus, was observed at later time points and was associated with the emergence of resistance. The majority of resistant substitutions observed in vivo were at residues 28, 30, 31, and 93 for genotype 1a and at residues 31 and 93 for genotype 1b. This is consistent with the resistance identified in vitro, confirming the utility of the replicon system for assessing levels of resistance in response to BMS-790052 exposure.
In general, the single amino acid substitutions and some double amino acid substitutions (Q54H-Y93H) that were observed in genotype 1b conferred minimal resistance (1- to 28-fold; Table 1); however, some double amino acid substitutions in genotype 1b, such as L31V-Y93H, conferred high levels of resistance (14,789-fold; Table 1). For genotype 1a, the major resistance substitutions observed in the study delivered greater resistance than single substitutions in genotype 1b (Table 2). Some of them displayed poor replicative ability, especially variants with substitutions at residue 93. However, other variants, such as Q30E and L31V, replicated as well as the wild-type replicon. Some variants with double amino acid substitutions (such as Q30R-H58D) also conferred high levels of resistance in the transient replication assays (Table 2). This can, at least partially, explain why viral breakthrough was more commonly observed in patients infected with genotype 1a than those infected with genotype 1b.
The frequencies of substitutions conferring resistance to BMS-790052 in the NS5A region (residues 28-32 and 93) were examined in sequences deposited in the European HCV database (http://euhcvdb.ibcp.fr/euHCVdb/, accessed on March 23, 2010).5 In this study, the variant with Q30R-H58D substitutions displayed the highest level of resistance (>400,000-fold) (Table 2). Though Q30R is a common genotype 1a BMS-790052-resistant mutation,5, 6 the H58D substitution was not detected in the European HCV database. However, H58D was also identified in patient Q who, like patient S, was a member of the 100-mg cohort. H58D was detected at day 4 in patient Q, but not at any other time points (Table 3E), suggesting that resistant variants can emerge from different paths under similar selective pressures.
Previously characterized resistant variants were detected in only 3 of 24 patients' (M, T, and V) baseline specimens by population sequencing. Patient M (60-mg cohort) was infected with genotype 1a virus. A Q30R substitution was detected at a level of ~10% at baseline, but reached ~60% at day 4. This profile suggests that the Q30R variant was suppressed by the 60-mg dose of BMS-790052, with a slower rate of decline than wild-type virus at day 1. Replacement of the Q30R variant with a Q30H-Y93H variant at day 14 suggests that this double amino acid substitution variant with a high level of resistance (EC50 value: 409.8 ng/mL or 553 nM; Table 2) was selected. Selection of this highly resistant linked variant could explain the viral breakthrough observed in patient M during treatment with 60 mg of BMS-790052. Patient T in the 100-mg cohort was infected with genotype 1b virus with baseline resistance (Q54H-Y93H). Consistent with the in vitro resistance profile of this variant (Table 1), patient T experienced a maximal HCV RNA decline of ~4.5 log10 at day 7, suggesting that the variant with a Q54H-Y93H substitution (100%) was initially suppressed by BMS-790052. The EC50 value of L31V-Q54H-Y93H, the dominant variant at day 14, was 36.1 ng/mL or ~49 nM. Trough concentrations of BMS-790052 in patient T were 153-546 ng/mL or 207-737 nM (data not shown), much higher than the EC50 value for L31V-Q54H-Y93H. Experiments to determine why the virus was not suppressed in patient T are ongoing. Patient V in the 30-mg twice-daily cohort was also infected with genotype 1a virus. A Q30H-Y93H variant, with a high level of resistance to BMS-790052, was detected at baseline (~100% of Q30H and ~45% of Y93H; Table 3F). It is likely that the presence of this variant at baseline accounts for the lack of viral suppression in patient V.
As we observed in the single-ascending dose study, significant HCV RNA decline was required to detect resistance variants by population sequencing.4 This observation suggests that these variants were either present at very low levels at baseline or were initially inhibited by BMS-790052. Because variants, such as genotype 1a Q30H in patient R (100-mg cohort), were detected at 4 hours (the first time point) postdosing, it is likely that the Q30H variant preexisted at baseline. Clonal analysis of the baseline specimens could address this possibility.
From a virology point of view, the antiviral effect of a specific DAA is mainly determined by two factors: intrinsic potency and resistance barrier. Because of the exceptional potency of BMS-790052, patients generally experienced an initial sharp HCV RNA decline, indicative of the inhibition of wild-type virus. A slow second phase of viral decline or a slight viral rebound was observed at later time points, consistent with an accumulation of resistant variants and suggesting the adaptation or selection of resistant variants with enhanced fitness. The emergence of resistance suggests that BMS-790052, like NS3 protease inhibitors12 and NS5B polymerase allosteric inhibitors,13 may have a low genetic barrier for resistance. Only a single-nucleotide change (UAU or UAC to AAU or AAC) at residue 93 (Tyr to Asn) of genotype 1a NS5A is required for HCV to acquire clinical resistance to BMS-790052 (Table 2). Furthermore, through either accumulation or novel mutation, linked substitutions emerged, such as Q30R-H58D (patient S, 100-mg cohort; Table 3E), which conferred a high level of resistance.
Questions not addressed in the current study remain. For example, how common is the linkage of resistance substitutions? The possible linkage of two or more substitutions may only be recognized by population sequencing when the substitution for each residue is >50%. Clonal analysis will reveal the frequency of linkage, and phenotypic analysis of variants with linked substitutions will provide useful information about the level of resistance contributed by linked variants. In addition, the rate of decay of resistant variants after cessation of dosing is currently unknown; however, studies to address this are ongoing.
To maximize the anti-HCV response and minimize resistance, combination therapy, similar to current HIV treatment, could be used to enhance the resistance barrier. During combination therapy, variants with multiple substitutions, generally accompanied by reduced fitness, are required to generate clinical resistance. Our in vitro data show that BMS-790052-resistant variants remain fully sensitive to inhibitors of the NS3 protease and the NS5B polymerase,5 demonstrating the potential for effective combination treatment with DAAs that have different mechanisms of actions and targets.
In conclusion, plasma trough concentrations of BMS-790052 monotherapy at the dose range used for the MAD study were not sufficient to prevent all viral breakthroughs because of the emergence of resistant variants. Because BMS-790052 is a novel class of HCV inhibitor with a demonstrated antiviral response in genotype 1-infected patients, it is anticipated that BMS-790052 will be an excellent candidate for combination therapy with interferon plus ribavirin and/or other small-molecule HCV inhibitors. It is also anticipated that combination therapy will suppress the selection of resistant variants.
|
|
| |
| |
|
|
|