 |
|
|
| |
Combinations of Primary and Secondary Integrase Mutations in the VIKING Pilot Study: Effects on and Rationale for Dolutegravir Dissociation
|
| |
| |
Reported by Jules Levin
International Workshop on HIV & Hepatitis Virus Drug Resistance and Curative Strategies; June 5-9, 2012; Sitges, Spain
Kendra Hightower, Felix DeAnda, Ruolan Wang, Cindy Vavro, Mark Underwood
GlaxoSmithKline, Research Triangle Park, NC, USA

Abstract
BACKGROUND: VIKING is a phase IIb clinical trial of the integrase inhibitor (INI) dolutegravir (DTG, S/GSK1349572) for INI-experienced subjects harboring raltegravir (RAL)-resistant HIV-1. To gain a better understanding of clinical responses based on integrase mutation
accumulation and combinations, INI binding and molecular modeling studies were conducted with integrase (IN) proteins identified from VIKING; these included multiple secondary and RAL-associated signature resistance mutations.
METHODS: Multiple substitutions were introduced into wild-type HIV-1 integrase based on population genotypes observed during VIKING. Dissociation of [3H]-labeled DTG, RAL, or elvitegravir (EVG) from integrase/DNA/bead complexes was monitored using unlabeled compound. Structural models of HIV-1 integrase/DNA, both wild-type and with select amino acid substitutions, were constructed from available crystallographic data on HIV-1 integrase and wild-type and mutant prototype foamy virus intasomes.
RESULTS:
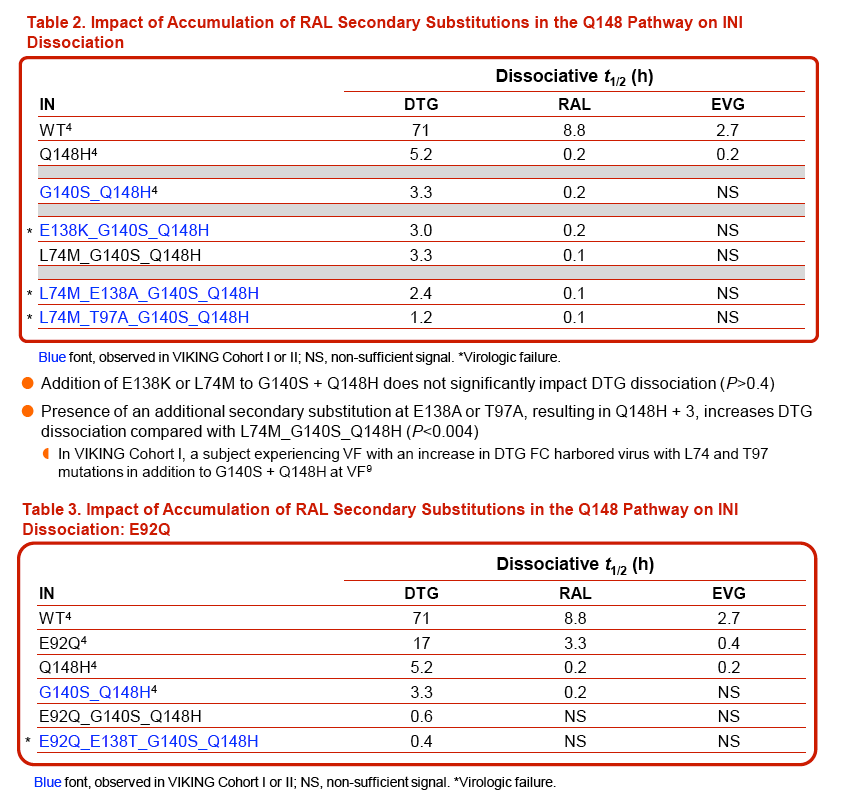
The addition of Y143H to G140S + Q148H integrase was not detrimental for DTG binding (dissociative t1/2 of 3.3 h for G140S + Q148H vs 3.9 h + Y143H).
DTG had a half-life of 2.3 h with integrase having substitutions at Y143R + N155H in conjunction with T97A; no impact on DTG dissociation was observed with additional substitutions at L74M or L74M + E138A.
For G140S + Q148H, addition of a L74M or E138K substitution did not impact DTG dissociation relative to G140S + Q148H (t1/2 of 3-3.3 h vs 3.3 h) but the accumulation of further substitutions (Q148H + 3) at L74M, T97A, or E138A did significantly impact DTG dissociation.
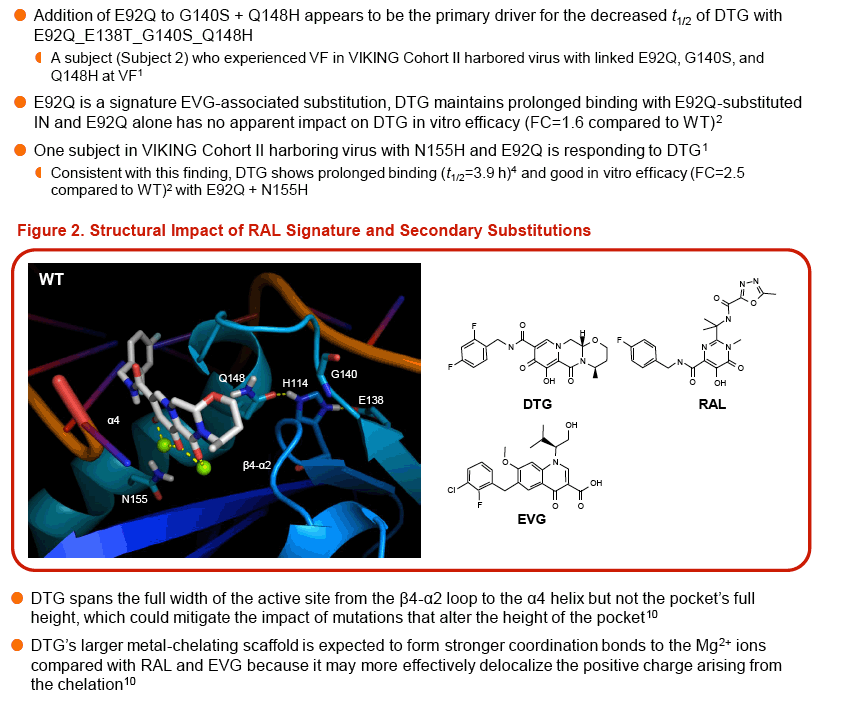
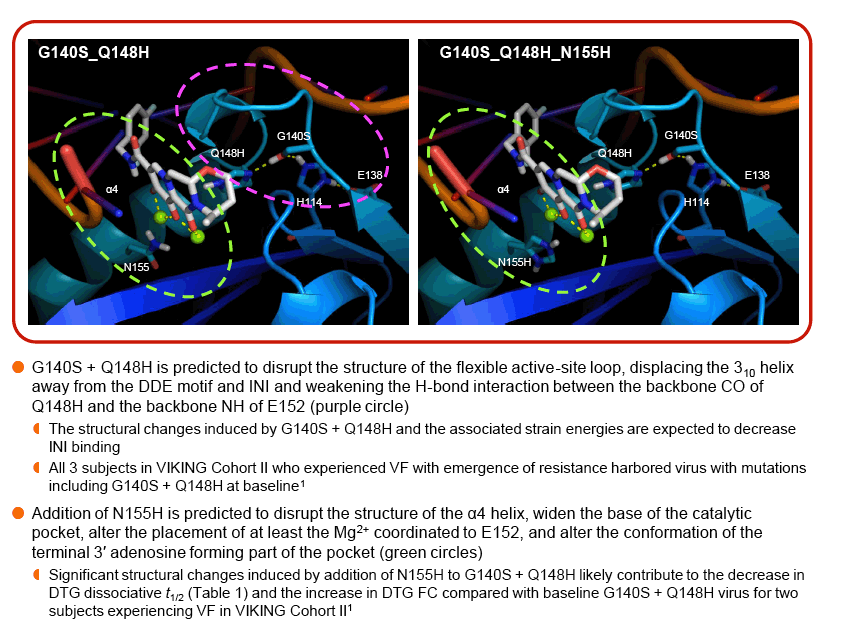
Modeling suggests that E138A/K substitutions may play a role in disrupting the active-site loop, while the L74M and T97A substitutions may impact the configuration of the active site and the placement of the metal coordination complex.
With all of the multi-substituted integrases, dissociation of RAL and EVG was fast (t1/2<1 h) or binding was insufficient for analysis.
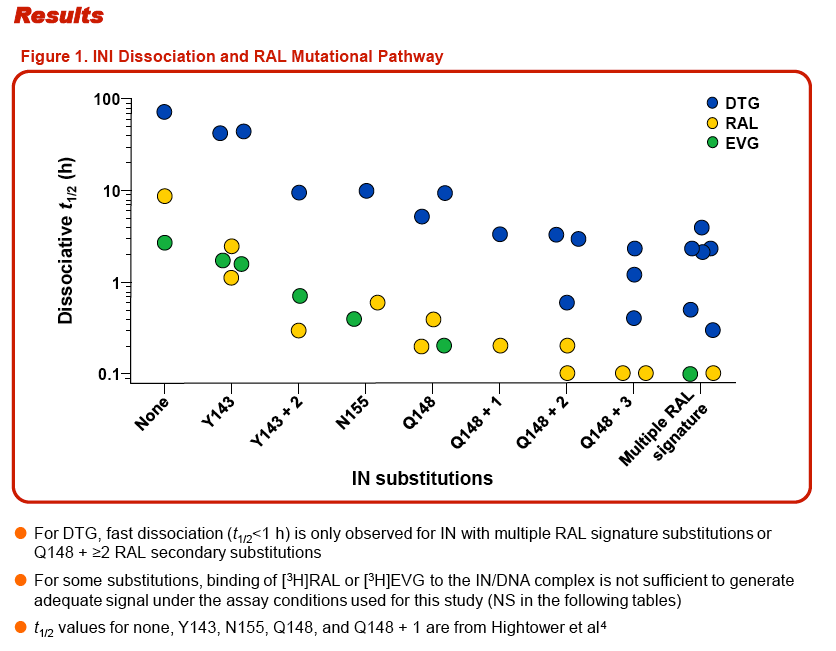
CONCLUSIONS: The presence of multiple integrase substitutions could cause significant disruption in the INI binding pocket thereby impacting development of INI resistance. The protracted DTG binding (t1/2>1 h) observed with integrase/DNA complexes containing several RAL-associated signature and secondary integrase substitutions suggests that DTG can tolerate greater disruption in the integrase active site than RAL and EVG and that the accumulation of multiple RAL-associated mutations is required to break through DTG's barrier to resistance.

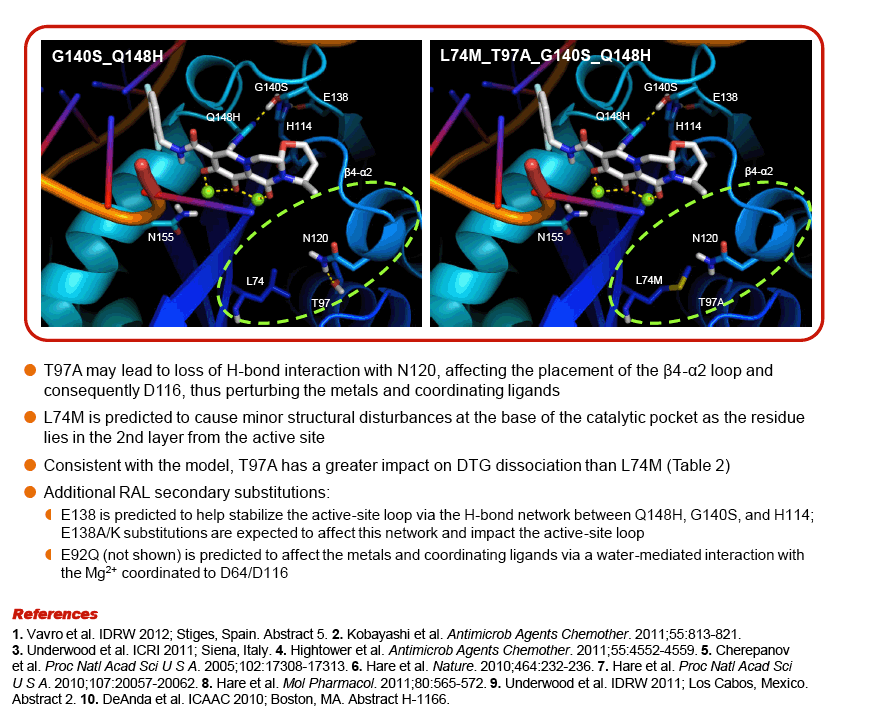
|
| |
|
|
|
|
|