 |
|
|
| |
Greater Bone Deficits Seen in Young US Men Infected at Birth Versus in Adolescence....bone senescence
|
| |
| |
CROI 2014, March 3-6, 2014, Boston
Mark Mascolini
Compared with young HIV-negative US men, young men infected with HIV perinatally or during adolescence had lower peak bone mass and fewer circulating osteogenic precursors, which give rise to functional osteoblasts (bone-forming cells) [1]. And the perinatally infected group had greater bone deficits than the group infected during adolescence. These findings from a cross-sectional comparison of perinatally infected men, men infected during adolescence, and HIV-negative men point to the possibility of greater long-term bone loss and higher fracture rates in people with HIV.
Michael Yin and Columbia University colleagues noted that peak bone mass occurs in a person's 20s. If peak bone mass is lower in people infected with HIV during adolescence and lower still in those infected perinatally, they may be expected to have weaker bones and a higher fracture risk at an earlier age than people without HIV.
Earlier research by these investigators found that men infected with HIV at an early age have 6% to 19% lower volumetric bone mineral density (BMD), markedly abnormal trabecular plate and cortical microarchitecture, and decreased whole bone stiffness (a measure of bone strength) by high-resolution peripheral quantitative CT (HRpQCT) of the radius and tibia [2]. Inadequate bone formation, the researchers noted, may result from insufficient bone formation (by osteoblasts) or excessive bone resorption (by osteoclasts).
Continuing to explore these issues, Yin and coworkers asked whether chronic immune activation with HIV infection promotes accelerated replicative senescence of osteoblast precursors. The study involved 58 HIV-positive and negative men from 20 to 25 years old, the period of peak bone mass. All HIV-positive men had taken antiretroviral therapy for more than 1 year. The Columbia team used DXA scans and HRpQCT to compare peak bone mass, microarchitecture, and bone strength in 18 perinatally infected men, 20 men infected during adolescence, and 20 uninfected men. By flow cytometry they compared numbers of circulating osteogenic precursors in these three groups. They measured osteogenic precursor telomere length (a signal of cell senescence) with quantitative PCR.
The perinatal, adolescent, and HIV-negative groups averaged 22, 23, and 21 years in age (P < 0.05 when comparing the perinatal and adolescent groups or the HIV group with the HIV-negative group). Respective black/Hispanic proportions were 61%/39%, 50%/50%, and 40%/60%. Body mass index averaged 25 or 26 kg/m(2) across the three groups.
Among perinatally infected, current and nadir CD4 counts averaged 505 and 181, while men infected in adolescence had current and nadir averages of 531 and 292. The perinatal group had taken antiretrovirals for an average 12 years versus 2 years in the adolescent group, and a higher proportion of the adolescent group (65% versus 11%) had taken a protease inhibitor. Most men in both groups had taken tenofovir. A lower proportion in the perinatal group than the adolescent group had a viral load below 20 copies (56% versus 65%).
Bone mineral density (BMD) was significantly lower (by 5% to 10%) in HIV-positive than negative men at the femoral neck, total hip, distal radius, and ultra distal radius--and nonsignificantly lower at the lumbar spine (P = 0.09). At the radius and tibia, HRpQCT documented 8% to 18% lower total volumetric BMD, trabecular volumetric BMD, trabecular thickness, cortical thickness, and stiffness in HIV-positive men. Comparing BMD and bone stiffness in perinatally infected and adolescently infected men with HIV, Yin and colleagues found significantly lower BMD in the ultra distal radius of the perinatal group (P = 0.02) and 14% lower bone stiffness at the radius in perinatally infected men (P = 0.02).
Numbers of circulating osteogenic precursors were significantly lower in HIV-positive than negative men (P < 0.05) and significantly lower in the perinatal group than the adolescent group of men with HIV (P < 0.05). Osteogenic precursors were significantly lower in HIV-positive than negative men when the researchers limited the analysis to HIV-positive men with an undetectable viral load. The telomere-to-single copy gene ratio of osteogenic precursors was significantly lower in men with than without HIV and significantly lower in the perinatal group than the adolescent group (P < 0.05 for both comparisons).
Proportions of circulating osteogenic precursors in HIV-positive and negative men correlated positively with radial bone stiffness when the researchers characterized those cells as LIN-OCN+/RUNX2+ (r = 0.277, P = 0.037) or LIN-OCN+/RUNX2+CD34- (r = 0.368, P = 0.005). The correlation is stronger in CD34- cells, which are thought to be the more mature osteogenic precursor population.
Yin and colleagues concluded that young men infected with HIV perinatally or during adolescence had (1) lower peak bone mass, (2) lower bone strength by stiffness, and (3) lower numbers of circulating osteogenic precursors with short telomere lengths than similarly aged men without HIV. These deficits proved greater in perinatally infected men than in men infected during adolescence.
The researchers proposed that "HIV infection early in life may result in replicative senescence of osteoblast precursors which may lead to decreased numbers of functional osteoblasts." As a result, men infected with HIV early in life may face impaired bone acquisition during adolescence, which could lead to decreased osteoblast reserves in older age.
References
1. Manavalan JS, Arapdi S, Shah J, et al. Replicative senescence of circulating osteogenic cells and low BMD in perinatally infected men. CROI 2014. Conference on Retroviruses and Opportunistic Infections. March 3-6, 2014. Boston. Abstract 132. http://www.croiwebcasts.org/console/player/22266?mediaType=slideVideo&
2. Yin MT, Lund E, Shah J, et al. Lower peak bone mass and abnormal trabecular and cortical microarchitecture in young men infected with HIV early in life. AIDS. 2013 Sep 25. Epub ahead of print.
-----------------------------------

Whether these observed differences in bone strength by imaging will result in early fractures among in men infected with HIV early in life is uncertain, but the magnitude of difference in stiffness is similar to the 10-20% difference seen in postmenopausal women with and without fracture, and is therefore clinically relevant.
Taken together our bone cell data, leads us to speculate that HIV infection early in life may result in replicative senescence of osteoblast precursors which may lead to decreased numbers of functional osteoblasts and result in impaired bone acquisition.
We have studies planned to evaluate the association between immunological parameters and osteogenic precursors, as well as characterizing the functional characteristics these circulating osteogenic prescursors

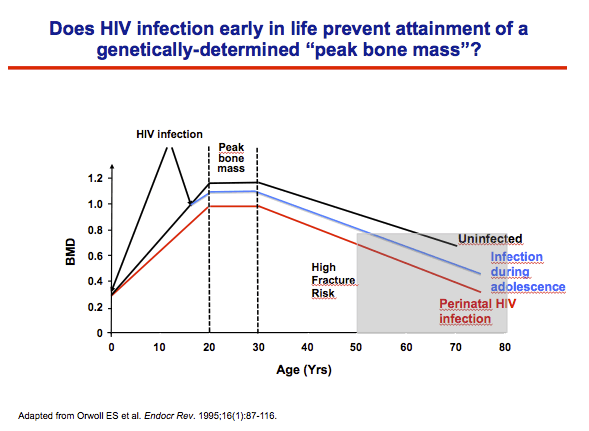
In development, a genetically determined peak bone mass is attained in both men and women in their early 20�s; after which there is a steady decline in bone mass and bone strength until late adulthood when fracture risks increase
HIV infection and ART are both associated with bone loss.
We hypothesized that HIV infection that occurs early in life would prevent an individual from attaining their genetically-determined Peak Bone mass, resulting in a lower starting point for decline in bone mass in adulthood and potentially translate to fractures at an earlier age.
This figure depicts the hypothetical trajectory of bone acquisition with early HIV infection, with perintally-infected individuals attaining lower PBM than either uninfected individuals or individuals infected during adolescence

inadequate bone acquisition may result from either insufficient bone formation as a result of decreased numbers or function of osteoblasts or excessive bone resorption as a result of increased numbers or activity of osteoclasts
While most researchers attribute bone loss with HIV to increased bone resorption caused by inflammatory cytokines,
HIV infection may also have a negative effect on bone formation,
This may be particularly important in children and adolescents who are still growing
in vitro studies suggest that HIV might have direct or indirect effects on osteoblasts causing apoptosis or decreased differentiation from mesynchymal cells precursors.
Another potential mechanism is suggested from a finding in Rheumatoid arthritis, another disease of chronic inflammation. In a biopsy study, periarticular osteoblasts from rheumatoid arthritis patients were found to have a more senescent phenotype with shorter telomere length and decreased replicative lifespan in culture than periarticular osteoblasts from patients with osteoarthritis.
Osteoblast senescence is a recognized contributor to osteoporosis of aging.
Therefore, could chronic immune activation with HIV also promote accelerated replicative senescence of osteoblast precursors?
Alternative text:
If accelerated periarticular osteoblast senescence is a consequence of the chronic inflammatory process of rheumatoid arthritis, chronic immune activation with HIV infection could also result in accelerated osteoblast senescence in the bone microenvironment.
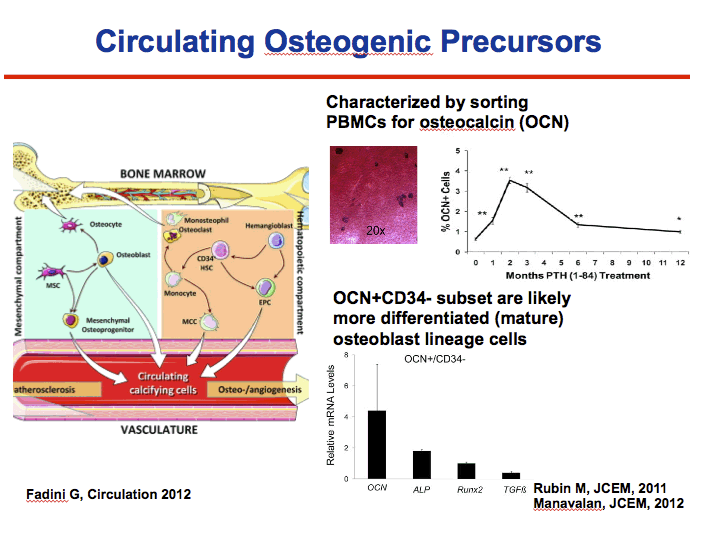
It is hard to get bone biopsies on patients to prove that point
However, it is increasingly accepted that osteoblast precursors also circulate in blood vessels.
In this review article by Fadini and Khosla, they summarize data that supports the presence of circulating calcifying cells that arise from mesenchymal or hematopoietic lineages from the bone marrow and traffic through blood stream either to the bone to repair fracture or form bone, or to the endothelium to calcify atherosclerotic lesions
Exactly which cellular markers define the subpopulation that are specifically home to the bone as opposed the the endothelium is still controversial
Ostecalcin is an osteoblast specific protein
by using flow cytometry to sort for PBMCs with an osteocalcin antibody Drs Manavalan and Kousteni who are bone biologists at Columbia University have defined a population of cells that behave like osteoblast.
In culture, these cells form mineralizing nodules. In addition, in hypoPTH patients who received PTH treatment, numbers of OCN+ cells increased in their periphery and correlated with indices of increased bone formation on bone biopsies
In addition, they have defined a subpopulation of OCN+ cells that have lost the hematopoietic CD34 marker as more differentiated or mature osteoblast lineage cells, since they express greater levels of RUNX2, a major transcriptional factor of osteoblasts, than CD34+ cells.

In order to determine the effect of HIV infection on peak bone mass and osteoblasts, We performed a cross sectional study 58 HIV+ and HIV- african amerian and hispanic men between ages of 20-25, an age when they should be at peak bone mass.
HIV infected men were either perintally-infected or infected during adolescence and had to have been on ARVs of any type for at least 1 year
The FIRST aim was to compare peak bone mass, microarchitecure and bone strength within the 3 groups by DXA and HRpQCT of the radius and tibia which has high enough resolution to determine cortical and trabecular indicies. In addition, we measured stiffness using FEA, which is a validated simulated measure of true bone strength that correlates with Fracture
The SECOND aim was to compare the numbers of circulating osteogenic cells between the 3 groups and correlate those findings with bone indices
We refined the original methods by first sorting for LIN- cells to increase the precursor and stem cell population and then sorting by osteocalcin and labeling for RUNX2, a key transcriptional factor for osteoblast differentiation, to increase specificity. We also used a lack of CD34 as a marker for more mature osteogenic cells.
Lastly, we used quantitiatve PCR to measure telomere lengths of osteocalcin+ cells.

The SECOND aim was to compare the numbers of COP cells and correlate those findings with bone indices
We refined the original methods using flow cytometry by first gating for lineage negative (LIN-) cells to increase the precursor and stem cell population and then gating on osteocalcin and intracellular RUNX2, a key transcriptional factor for osteoblast differentiation to increase the specificity for an osteogenic cell. As before, we also used the lack of expression of CD34 to define more mature osteogenic cells.
Lastly, we used quantitiatve PCR to measure telomere lengths of flow cytometry sorted Lineage negative, osteocalcin + cells.
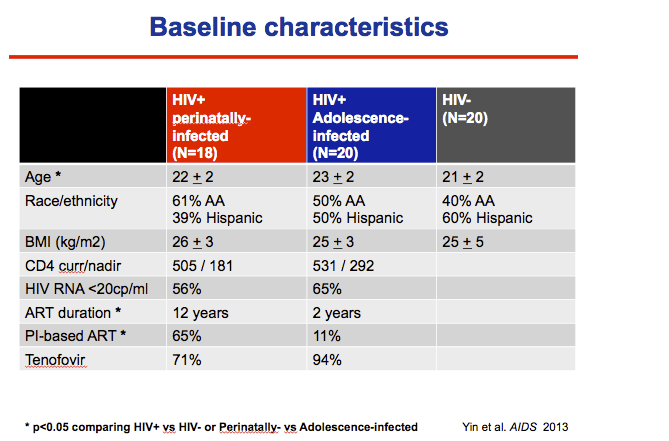
the perintally infected group had longer ART exposure and more frequent PI use, Tenofovir use did not differ between groups.
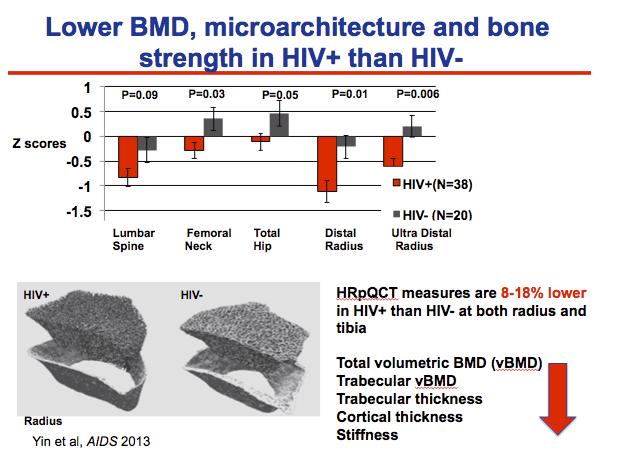
In this figure, we compare the areal BMD by Z scores, which adjusts for age and race, at the spine, hip and radius,
HIV infected men (combining both perinatal and adolescent infected groups) had 5-7% lower BMD at the hip and radius than uninfected controls,
In the CT image of the radius on the bottom left of the HIV infected individual appears to have a thinner cortex, on the outer shell, and thinner trabeculae the middle
Comparing the actual measures, the HIV+ group had lower total and volumetric BMD, and lower, trabecular and cortical thickness and stiffness at both the radius and tibia than the uninfected group
The magnitude of this difference was between 8-18%
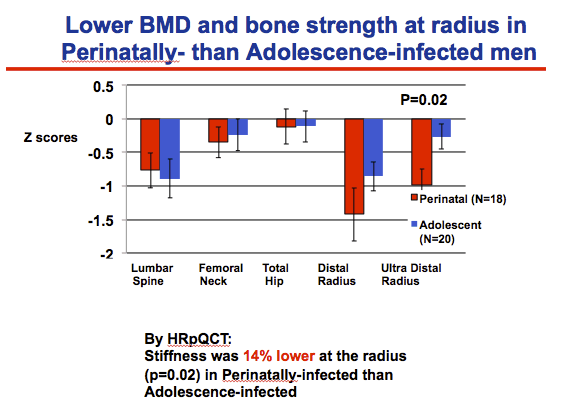
Differences between the Perinatally-infected and adolescence infected groups were less pervasive.
The perinatal-infected group had lower BMD by DXA than the adolescence-infected group ONLY at the ultradistal radius.
This finding was supported by the HRPQCT, which found that stiffness was 14% lower at the radius in perinatally-infected than adolescence-infected groups.
Stiffness 19% lower at the tibia (p=0.052)
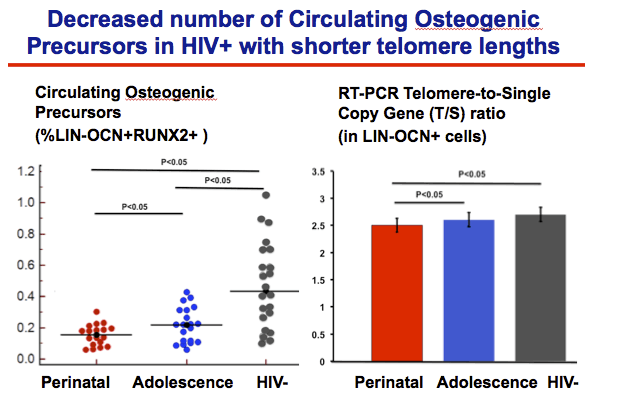
Next, using flow cytometry we compared the total number COP cells compared between 3 groups , again these are the LIN-OCN+ RUNX2+.
These cells represent <1% of total PBMCs
we found that the proportion of PBMCs representing circulating osteogenic progenitor cells (LIN-/OCN+/Runx2+) were lower in either HIV-infected groups than uninfected controls.
Perinatally-infected group had the lowest number, lower than both the adolescent infected and the uninfected,
Next we measured the telomere lengths of the LIN-OCN+ cells using QPCR, estimated by the Telomere to single copy Gene ratio,
The telomere lengths were shorter in the perintally infected group than both the adolescent-infected and the uninfected cpntrols.
CD34 cells did not differ between HIV+ and HIV-
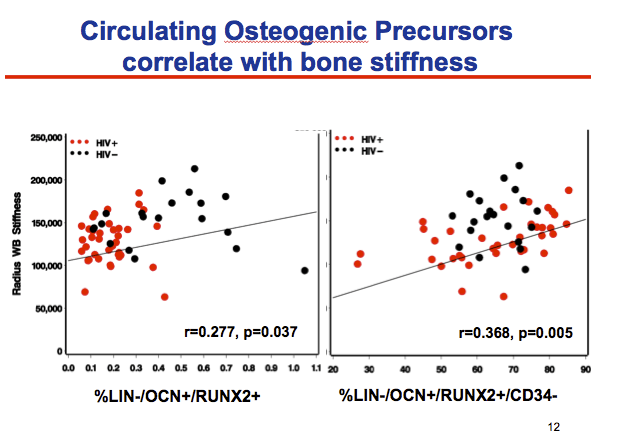
Last, we evaluated the correlation between ciculating osteogenic precursors and the stiffness index
we found a modest correlation between higher number of OCN+RUNX2+ cells and higher whole bone stiffness at the radius
When we looked at the the CD34- subpopulation, which are postulated to be the more mature circulating osteogenic precursors, we found a slightly higher correlation with radial bone stiffness.
This suggests that the mature osteogenic cells may be more representative of (1) either the population that can home from the circulation to the bone or (2) just better reflect the quantity of functional osteoblasts in the bone.
|
| |
|
|
|
|
|