| |
GS-9620, an Oral Agonist of Toll-Like Receptor-7,
Induces Prolonged Suppression of Hepatitis B Virus in Chronically Infected Chimpanzees
|
| |
| |
Download the PDF
"Short-term oral administration of GS-9620 provided long-term suppression of serum and liver HBV DNA. The mean maximum reduction of viral DNA was 2.2 logs, which occurred within 1 week of the end of GS-9620 administration; reductions of >1 log persisted for months. Serum levels of HBV surface antigen and HBV e antigen, and numbers of HBV antigen-positive hepatocytes, were reduced as hepatocyte apoptosis increased. GS-9620 administration induced production of interferon-α and other cytokines and chemokines, and activated interferon-stimulated genes, natural killer cells, and lymphocyte subsets. The small molecule GS-9620 activates Toll-like receptor 7 signaling in immune cells of chimpanzees to induce clearance of HBV-infected cells. This reagent might be developed for treatment of patients with chronic HBV infection."
"In this study, only 3 HBV chronically infected animals were available for the project; all were chronically infected for more than 2 decades. This limited the scope of the study to proof-of-concept that TLR-7 stimulation could impact HBV chronic infection. We were not able to evaluate regimen optimization or combination with direct antiviral therapy or to determine whether extended dosing at the low dose or combination with antiviral agents could result in the same reduction in viral DNA and antigen-positive hepatocytes without an increase in liver enzymes. In this context, it is also important to note that the results of this study presented evidence for a "presystemic" TLR-7 response: low doses of GS-9620 were capable of inducing an antiviral immune response at the level of the liver, PBMC, and possibly gut-associated lymphoid tissue without induction of systemic IFN-α or its side effects. At the low dose, in both uninfected and infected animals, GS-9620 induced ISGs in the liver and/or in PBMC with no detectable increase in serum levels of IFN-α. Ongoing clinical trials are evaluating whether administration of GS-9620 at presystemic dose levels provides similar antiviral benefits alone, as recognized in this study, or in combination with direct-acting antivirals, without the side effects often associated with systemic administration of IFN-α. Identification of a well-tolerated, finite therapeutic regimen, including the possible combination of immune modulators, such as GS-9620, with direct-acting antiviral agents for the treatment and cure of chronic HBV infection in a significant and clinically relevant percentage of treated patients would be transformative for this disease."
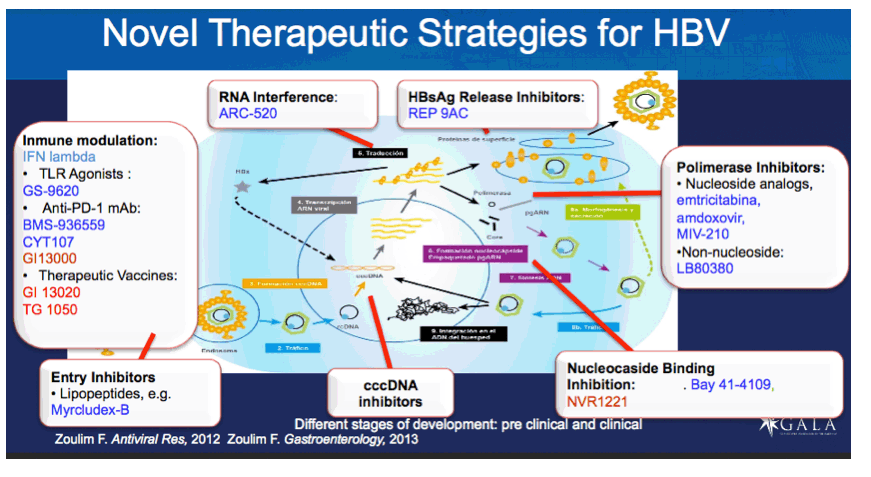
"The ultimate goal of therapy for HBV chronic infection is viral eradication and cure of the underlying liver disease.22 The greatest advances in therapy have been made with nucleos(t)ide analogues that are chain terminators of the reverse transcription process.1, 23 Although nucleos(t)ide therapies reduce circulating virus to undetectable levels,24 they fail to eliminate infected hepatocytes, primarily due to the inability of this approach to eradicate the nonreplicating and stable form of viral DNA in the nucleus, covalently closed circular DNA. Although viral resistance was a major issue with first-generation nucleos(t)ide reverse transcriptase inhibitors,3 newer analogues such as tenofovir appear to have little if any potential for the development of resistance during long-term therapy.25, 26 Nonetheless, the percentage of treated patients that develop loss of HBsAg is small, only 10% over 5 years with tenofovir and only 5% in 2 years with entecavir. Pegylated IFN-α therapy suppresses viremia to undetectable levels in only a small percentage of patients during 48 weeks of therapy, yet some patients exhibit apparent cure (no rebound of viremia off of therapy, loss of HBsAg, and seroconversion with detectable antibodies to HBsAg), and the percentage of patients experiencing cure increases for several years after cessation of therapy.4, 5, 6 These data suggest that the use of improved immunomodulators, such as the TLR-7 agonist GS-9620, can lead to a clinically relevant improvement in therapy, with a significantly higher incidence of viral eradication in patients with chronic HBV infection."
GS-9620, an Oral Agonist of Toll-Like Receptor-7, Induces Prolonged Suppression of Hepatitis B Virus in Chronically Infected Chimpanzees
Gastroenterology June 2013
ROBERT E. LANFORD,1,2,* BERNADETTE GUERRA,1 DEBORAH CHAVEZ,1 LUIS GIAVEDONI,1,2 VIDA L. HODARA,1
KATHLEEN M. BRASKY,2 ABIGAIL FOSDICK,3 CHRISTIAN R. FREY,4 JIM ZHENG,5 GRUSHENKA WOLFGANG,3
RANDALL L. HALCOMB,6 and DANIEL B. TUMAS3,4,*
1Department of Virology and Immunology, Texas Biomedical Research Institute, San Antonio, Texas; 2Southwest National Primate Research Center, San Antonio, Texas; and Departments of 3Drug Safety Evaluation, 4Biology, 5Drug Metabolism, and 6Medicinal Chemistry, Gilead Sciences, Inc, Foster City, California
Background & Aims
Direct-acting antiviral agents suppress hepatitis B virus (HBV) load, but they require life-long use. Stimulation of the innate immune system could increase its ability to control the virus and have long-lasting effects after a finite regimen. We investigated the effects of immune activation with GS-9620Ña potent and selective orally active small molecule agonist of Toll-like receptor 7Ñin chimpanzees with chronic HBV infection.
Methods
GS-9620 was administered to chimpanzees every other day (3 times each week) for 4 weeks at 1 mg/kg and, after a 1-week rest, for 4 weeks at 2 mg/kg. We measured viral load in plasma and liver samples, the pharmacokinetics of GS-9620, and the following pharmacodynamics parameters: interferon-stimulated gene expression, cytokine and chemokine levels, lymphocyte and natural killer cell activation, and viral antigen expression. Clinical pathology parameters were monitored to determine the safety and tolerability of GS-9620.
Results
Short-term oral administration of GS-9620 provided long-term suppression of serum and liver HBV DNA. The mean maximum reduction of viral DNA was 2.2 logs, which occurred within 1 week of the end of GS-9620 administration; reductions of >1 log persisted for months. Serum levels of HBV surface antigen and HBV e antigen, and numbers of HBV antigen-positive hepatocytes, were reduced as hepatocyte apoptosis increased. GS-9620 administration induced production of interferon-α and other cytokines and chemokines, and activated interferon-stimulated genes, natural killer cells, and lymphocyte subsets.
Conclusions
The small molecule GS-9620 activates Toll-like receptor 7 signaling in immune cells of chimpanzees to induce clearance of HBV-infected cells. This reagent might be developed for treatment of patients with chronic HBV infection.
Therapeutic treatment of chronic hepatitis B virus (HBV) infection is currently limited to nucleos(t)ide analogues and pegylated interferon-(IFN)-α.1, 2 First-line therapy for HBV is limited to the 2 nucleos(t)ide analogues tenofovir and entecavir, which are highly effective at suppressing viral replication and can reduce serum viral load to undetectable levels. However, these agents do not lead to viral eradication, potentially requiring life-long use and possible emergence of resistance.3 The potential for therapeutic immune modulation to treat HBV chronic infection is illustrated by durable responses, normalization of alanine aminotransferase (ALT), and sustained reduction in viremia attained in a small percentage (<20%) of patients treated for 1 year with pegylated IFN-α.4, 5, 6 A key observation is that the apparent cure rate after long-term high-dose IFN-α treatment increases for several years after treatment, based on loss of HBV surface antigen (HBsAg) and seroconversion for anti-HBsAg antibody. This supports the hypothesis that viral control is due to immune modulation and slow induction of a protective antiviral immune response. The low rate of HBsAg loss and seroconversion with current therapies illustrates the need for new approaches to induce a protective antiviral immune response and durable cure in patients with chronic HBV.
Toll-like receptor (TLR) 7 is a pathogen recognition receptor predominantly expressed in lysosomal/endosomal compartments of plasmacytoid dendritic cells (pDCs) and B lymphocytes that recognizes a pathogen-associated molecular pattern in viral single-stranded RNA.7 Upon stimulation of TLR-7, pDCs produce IFN-α8, 9 and other cytokines/chemokines and cause activation of natural killer (NK) cells and cross-priming of cytotoxic lymphocytes,10 thereby orchestrating both innate and adaptive immune responses.11 For these reasons, TLR-7 has been pursued as a therapeutic target for cancer, viral infections, and other diseases.12, 13, 14, 15 GS-9620 is a potent, orally active TLR-7 agonist with selectivity for induction of IFN-α over proinflammatory cytokines. Here, we demonstrate that a TLR-7 agonist provides therapeutic efficacy for treatment of HBV chronic infection in chimpanzees, the only primate model of persistent HBV infection.16, 17 The immune modulation induced by activation of TLR-7 resulted in rapid reduction of viremia, reduction in serum HBsAg and HBV e antigen (HBeAg) levels, and an apparent reduction of the numbers of infected hepatocytes with short-term therapy, and provided prolonged suppression of viremia after termination of therapy.
Results
Pharmacokinetics and Pharmacodynamics of GS-9620 in Uninfected Chimpanzees: Induction of IFN Response and Cytokines-Chemokines by TLR-7 Agonist GS-9620
GS-9620, a potent selective TLR-7 agonist, was designed to have rapid clearance and low-level systemic exposure after oral administration to allow for transient TLR-7 stimulation. Consistent with the selectivity of GS-9620 and the biology of TLR-7, chimpanzee PBMCs stimulated in vitro with GS-9620 displayed a lower minimum effective concentration for IFN-α, chemokines CXCL10 (IP-10), CCL7 (monocyte chemotactic protein [MCP]-3), and CCL4 (macrophage inflammatory protein [MIP]-1ß), interleukin (IL)-1 receptor antagonist, and IFN-γ in comparison to proinflammatory cytokines (Supplementary Table 1). In vivo, single, oral doses of GS-9620 at 0.3 and 1 mg/kg in uninfected chimpanzees demonstrated a dose- and exposure-related induction of serum IFN-α, select cytokines/chemokines, and ISG in the peripheral blood and liver. After oral administration at 0.3 mg/kg (n = 3) and 1 mg/kg (n = 3 and n = 4), GS-9620 Cmax was 3.6 ± 3.5 nM, 36.8 ± 34.5 nM, and 55.4 ± 81.0 nM, respectively. Peak serum interferon responses occurred at 8 hours post dose and are shown in Figure 1. Mean peak levels of induced serum IFN-α were 66 pg/mL and 479 pg/mL at doses of 0.3 mg/kg and 1 mg/kg, respectively (Figure 1). GS-9620 treatment induced ISG transcripts, including ISG15, OAS1 MX1, IP-10, and I-TAC in PBMC (Supplementary Table 2) at 0.3 mg/kg and in both PBMC and the liver at 1 mg/kg (Figure 2 and Supplementary Table 2). Serum levels of 42 different cytokines were evaluated. The magnitude and breadth of cytokine induction correlated with GS-9620 dose (Figure 2 and Supplementary Table 3). The 0.3-mg/kg dose induced ≥3-fold increases in serum IL-7, IL-10, IP-10, fractalkine, IL-1α, IL-1 receptor antagonist, and granulocyte colony-stimulating factor, whereas the 1-mg/kg dose induced ≥3-fold increases in the same cytokines (except IL-7) and serum IL-12p40, IL-12p70, MCP-1, MCP-3, MIP-1α, MIP-1ß, IL-8, IL-1ß, IL-6, tumor necrosis factor-ß, and neopterin. GS-9620 was well tolerated in uninfected chimpanzees; the only drug-related changes were transient increases in peripheral blood neutrophils and decreases in lymphocytes, consistent with cell trafficking induced by the cytokines and chemokines mentioned. Based on these data, 1 mg/kg was selected as the starting dose for treatment of HBV-infected animals.
Antiviral Efficacy of TLR-7 Agonist GS-9620 in HBV-Infected Chimpanzees
Therapeutic evaluation was performed in 3 chimpanzees that had chronic HBV infections for >24 years. One chimpanzee (4x0139) had high baseline serum HBV DNA, while the other 2 chimpanzees (4x0328 and 4x0506) had lower HBV DNA levels at baseline (Figure 3 and Supplementary Table 4). Serum levels of HBV DNA declined gradually in all 3 animals during the first treatment cycle, with a 1-log reduction in the high-titer animal (Figure 3A). The second treatment cycle caused a continued but more rapid decline of viral DNA in all 3 animals (Figure 3A-C), with a maximum viral reduction of 2.8 logs and a mean reduction of 2.2 logs (Figure 3 and Supplementary Table 5). Suppression of serum viral DNA levels by >1 log persisted for a minimum of 64 days. The viral load in the high-titer animal (animal 4x0139) was 1.8 logs below baseline at the end of the study, day 121, and remained >1 log below baseline for 280 days after initiation of dosing. The 2 low-viral-load animals returned to within 1 log of baseline within 100 and 71 days of the initiation of dosing, but continued to be suppressed by approximately 1 log for 1 to 2 years after this study.
Treatment also caused a decline in HBV viral DNA in the liver of the high-titer animal (animal 4x0139). The decline in liver HBV DNA paralleled the decline in serum DNA, 1.0 and 2.1 logs at the end of the first and second treatment cycles, respectively. The 2 low HBV DNA titer animals (animals 4x0328 and 4x0506) had very low levels of hepatic HBV DNA at baseline and did not exhibit a significant decline in viral DNA in the liver during therapy (data not shown). The apparent lack of decline in hepatic viral DNA might have been due to limitations in the assay and background in the assay imposed by the presence of integrated viral DNA.
HBsAg and HBeAg are secreted from HBV-infected hepatocytes independent of viral particles and are important clinical markers of infection independent of viral DNA levels. In the high-titer animal (4x0139), GS-9620 treatment reduced HBsAg and HBeAg serum levels by 61% and 93% from baseline, respectively (Figure 3C and D), and levels remained suppressed through post-treatment follow-up. Although the low-titer animals (4x0328 and 4x506) had low HBsAg levels at baseline, declines of 48% to 60% in HBsAg still occurred in both animals during therapy (Figure 3C). One of the low-titer animals (4x0328) was HBeAg positive at baseline and had a decline of 55% in HBeAg, while the other low-titer animal, 4x0506, was anti-HBe-positive at baseline (Supplementary Table 4). The rapid declines in liver viral DNA and secreted viral antigens in the high-titer animal are consistent with an elimination of infected cells, thus we directly examined the elimination of infected cells by immunohistochemical staining of liver sections for HBV core antigen (HBcAg). In the high-titer animal, approximately 30% of hepatocytes were positive for HBcAg staining before therapy (Figure 3E), and on the last day of dosing, when HBV DNA levels were reduced by >100-fold, few core-positive cells were detected (Figure 3F). These results are in stark contrast to those observed in patients during therapy with nucleos(t)ide analogues, which can reduce serum HBV DNA by ≥4 logs, yet no significant reduction occurs in serum HBsAg or HBcAg-positive hepatocytes over 48 weeks of therapy.20 Unfortunately, the number of HBcAg-positive cells was too low in the low-titer animals to accurately determine the degree of elimination.
Induction of Cytokines and Chemokines and ISGs by TLR-7 Agonist GS-9620 in HBV-Infected Chimpanzees
Levels of serum IFN-α and 38 other serum cytokines and chemokines were evaluated at pretreatment and at regular intervals during each treatment cycle. Prestudy IFN-α levels were below the limit of detection in animals 4x0139 and 4x0328, and these animals had dose-dependent increases in IFN-α after administration of GS-9620 at 1 mg/kg (mean, 119 pg/mL) and 2 mg/kg (mean, 700 pg/mL), although increases above baseline were not observed at every time point (Supplementary Table 6). The highest serum levels of IFN-α induced at the 2-mg/kg dose were 1396 pg/mL and 1545 pg/mL for animals 4x0139 and 4x0328, respectively (Supplementary Tables 7 and 8). The pretreatment baseline level of serum IFN-α was high in animal 4x0506 (1160 pg/mL) and was not further induced by GS-9620 treatment (Supplementary Table 9). This animal also had an elevated pretreatment baseline level of serum IFN-γ, yet GS-9620 treatment induced up to a 53-fold increase in serum levels of IP-10, a chemokine induced by IFN-α and IFN-γ. Of the other 38 cytokines and chemokines examined, during the first treatment cycle (1 mg/kg) only IL-10, MCP-3, and IL-1α were increased 5-fold above baseline, and during the second treatment cycle (2 mg/kg), 13 cytokines and chemokines were induced ≥5-fold; with IL-7, MIP-1ß, TNF-α, and granulocyte colony-stimulating factor being induced <10-fold; and IFN-α, IL-10, IP-10, MCP-1, MCP-3, IL-8, IL-1α, IL-1 receptor antagonist, and IL-6 being increased >10-fold (Supplementary Table 6).
Induction of ISG transcripts (ISG15, OAS1, MX1, IP-10, and I-TAC) was evaluated in PBMC and liver biopsy samples, and each was increased in both compartments in response to GS-9620 at both the 1- and 2-mg/kg dose levels; however, induction was not consistently present at the 1-mg/kg dose level for all days evaluated. ISG transcripts are rapidly up-regulated and down-regulated within a few hours of stimulation.21Variability in the level of response may be technical and related to the use of a single time point to measure a response that may be increasing or decreasing from the maximum value at the time of sampling (8 hours post dose), although exposure to GS-9620 may have varied to some extent after individual doses. Induction of ISGs was both more consistently present and the fold increases were greater at the 2-mg/kg dose (Figure 4 and Supplementary Tables 7, 8, 9, and 10). The group mean increase in transcript levels of the chemokine IP-10 in PBMC was 49.6- and 194-fold during the first (1 mg/kg) and second (2 mg/kg) treatment cycles, respectively (Figure 4 and Supplementary Tables 7, 8, 9, and 10). Interestingly, despite high pretreatment levels of serum IFN-α in animal 4x0506, and no apparent increase in IFN-α levels after GS-9620 administration, GS-9620 administration caused increases in ISGs in both PBMCs and the liver during both treatment cycles in this animal (Supplementary Table 10). Because TLR-7 induction in PBMCs by IFN-α was previously observed in chimpanzees,21 the induction of TLR-7 transcript was measured in this study. At pretreatment, the relative expression of TLR-7 in these chronically infected animals was 30-fold higher in PBMC than liver. TLR-7 levels were increased at multiple time points in the liver during treatment, with a mean maximum induction of 11.9-fold, while increases in PBMC were minimal at most time points, with a mean maximum induction 4.4-fold compared with pretreatment levels (Figure 5).
Activation of T Cells and NK Cells by TLR-7 GS-9620 Agonist in HBV-Infected Chimpanzees
Because the stimulation of TLR-7 in pDCs can result in the subsequent activation of immune effector cells, we evaluated the activation status of peripheral blood lymphoid and NK cell subsets using cell surface CD69 expression as a biomarker. During the second treatment cycle (2 mg/kg), an increase in the percentage of CD69-expressing CD8-positive T lymphocytes, NK, and NKT cells occurred, which was maximal after the first dose (Supplementary Figure 1). Mean fold increases in the percentage of CD69-positive cells ranged from 3.6 to 5.8 (Supplementary Table 11). No significant increases occurred during the first treatment cycle at 1 mg/kg (Supplementary Table 11).
Histological Changes in the Liver
In general, the severity of hepatic inflammation in chimpanzees associated with chronic HBV infection is less than that described in humans.18 A minimal to mild primarily lymphocytic inflammatory infiltrate in the portal tracts was present in all 3 animals before treatment. Changes noted during treatment included an increased mononuclear cell periportal infiltrate during the first treatment cycle, which, during the second treatment cycle, extended into adjacent hepatic parenchyma and sinusoids. Additionally, there was increased single-cell hepatocyte apoptosis, which was often associated with minimal clusters of mononuclear cells. Histological changes noted during treatment fully reversed within 3 to 5 weeks after treatment. Immunohistochemistry of biopsies at pretreatment and on the last day of therapy demonstrated a marked increase in hepatocellular expression of ISG15 protein, a marker of IFN-α induction (Figure 6), and an increased number of hepatocytes expressing activated caspase 3, a marker for apoptosis. The latter was associated with a correlative increase in hepatocellular regeneration and proliferation as determined by expression of Ki67.
Correlation of Viral Clearance and Elevation in Liver Enzymes
GS-9620 therapy was generally well tolerated, and no serious adverse events occurred during therapy. Clinical signs, body temperature, body weight, hematology, and blood chemistries were monitored throughout the study. Body weights were mildly decreased in all 3 animals during the study and recovered during the post-treatment period. Adverse events in the study were limited to anemia and transient increases in serum liver enzymes. Anemia was mild to moderate in all 3 animals, maximal reductions in red blood cell counts were 11% to 18%, and fully or partially recovered by the end of the study (day 121). Increases in serum levels of the liver enzymes ALT, aspartate aminotransferase, and γ-glutamyl transpeptidase (GGT) occurred during the second treatment cycle (2 mg/kg) (Figure 7). In the HBV high-titer animal (animal 4x0139), a sharp increase in the level of serum ALT and GGT occurred after the first week of treatment at 2 mg/kg, dosing was suspended for this animal for 1 week during which both ALT and GGT rapidly decreased, and then treatment resumed. No further increases in ALT were noted in this animal, however, GGT increases were noted (Figure 4). Liver enzyme elevations fully reversed after treatment; ALT and aspartate aminotransferase returned to pretreatment baseline levels within 3 weeks and GGT by the end of the study. Mild, transient 2- to 3-fold increases in serum total bilirubin occurred at single time points in 2 animals (day 43 in animal 4x0139 and day 57 in animal 4x0506) during treatment at 2 mg/kg and were concurrent with liver enzymes increases.
The transient and low-level single-incidence bilirubin increases were not considered adverse, but warrant monitoring in future clinical trials.
Discussion
The ultimate goal of therapy for HBV chronic infection is viral eradication and cure of the underlying liver disease.22 The greatest advances in therapy have been made with nucleos(t)ide analogues that are chain terminators of the reverse transcription process.1, 23 Although nucleos(t)ide therapies reduce circulating virus to undetectable levels,24 they fail to eliminate infected hepatocytes, primarily due to the inability of this approach to eradicate the nonreplicating and stable form of viral DNA in the nucleus, covalently closed circular DNA. Although viral resistance was a major issue with first-generation nucleos(t)ide reverse transcriptase inhibitors,3 newer analogues such as tenofovir appear to have little if any potential for the development of resistance during long-term therapy.25, 26 Nonetheless, the percentage of treated patients that develop loss of HBsAg is small, only 10% over 5 years with tenofovir and only 5% in 2 years with entecavir. Pegylated IFN-α therapy suppresses viremia to undetectable levels in only a small percentage of patients during 48 weeks of therapy, yet some patients exhibit apparent cure (no rebound of viremia off of therapy, loss of HBsAg, and seroconversion with detectable antibodies to HBsAg), and the percentage of patients experiencing cure increases for several years after cessation of therapy.4, 5, 6 These data suggest that the use of improved immunomodulators, such as the TLR-7 agonist GS-9620, can lead to a clinically relevant improvement in therapy, with a significantly higher incidence of viral eradication in patients with chronic HBV infection.
The mechanisms involved in viral clearance during acute and chronic HBV infection have been intensely examined, and although only partially understood, they are believed to be associated with antiviral CD8+ T cells trafficking to the liver, the production of IFN-γ, and the induction of immune inflammatory liver disease.16 The most challenging aspect of viral clearance with HBV is the elimination of nuclear covalently closed circular DNA, which is a nonreplicating DNA that can exhibit stability equal to the lifespan of the hepatocyte. Because 40%-100% of hepatocytes can be infected, a neutralizing antibody response to HBsAg is required to prevent infection of new hepatocytes that arise due to proliferation induced by hepatocyte death and/or uninfected cells arising from cytokine-induced noncytolytic elimination of the infection. Studies in transgenic mice have demonstrated that the innate immune response is capable of noncytolytic elimination of HBV replicative intermediates,27 and supportive studies in chimpanzees have observed a decrease in viral DNA before lymphocytic infiltration in the liver28, 29, 30 suggestive of noncytolytic mechanisms. However, studies in woodchucks during acute infection with woodchuck HBV have concluded that sufficient cell death occurs to account for viral clearance by cytolytic mechanisms, including loss of nuclear covalently closed circular DNA due to cell death and/or cell division.31 Collectively, these data suggest that both the innate and adaptive immune responses are critical for eradication of HBV infection.
This study demonstrated a long-term benefit from a short duration (8 weeks) of therapy with the oral TLR-7 agonist GS-9620. The mean maximal reduction in serum viral load was 2.2 logs, with a continued suppression of viral DNA by a minimum of 1 log for 2-4 months after dosing. Viral DNA remained suppressed in the high-titer animal for a period of 1 year, while both low-titer animals remained suppressed by approximately 1 log for at least 1 year. Consistent with stimulation of TLR-7 and activation of pDCs, the treatment induced select cytokines, including IFN-α, chemokines, and ISGs in PBMCs and liver. This response led to activation of specific populations of immune effector cells with increased expression of CD69 on CD8+ T and NK cells. Activation of pDCs is known to provide licensing for cross-priming of cytotoxic lymphocytes by classical licensing and NKT cell-dependent alternative licensing, and TLR-activated pDC recruit and activate NK cells to become more cytotoxic in vivo.10, 11, 32 GS-9620 induced a hepatic infiltration of mononuclear cells, including CD3+ and CD20+ lymphocytes, which, during the second treatment cycle, was also associated with single-cell hepatocellular apoptosis. The reduction in serum and liver HBV DNA was associated with a reduction in serum HBeAg, HBsAg, and HBcAg-positive hepatocytes and a concomitant increase in liver enzymes, hepatic immune cell infiltration, and hepatocellular apoptosis. Thus, the events occurring during therapy with GS-9620 are consistent with induction of antigen-specific T cell responses and NK cell responses with resultant selective killing of HBV-infected cells. The direct suppression of HBV replication by transient induction of type I IFNs and ISGs associated with TLR-7 activation might have contributed to the response, but would not account for the prolonged suppression of viral levels that was durable after cessation of treatment. The decline of serum HBsAg observed in all 3 animals and the apparent loss of core antigen-positive hepatocytes in the high-titer animal are not hallmarks of antiviral therapy with nucleos(t)ide therapy, and suggest a GS-9620 mechanism beyond suppression of viral replication. The results attained in this study are consistent with results we have reported in woodchucks chronically infected with woodchuck hepatitis virus, in which oral treatment with GS-9620 caused a sustained reduction in viral load, loss of woodchuck hepatitis virus surface antigen and seroconversion for antibodies to surface antigen.33 Collectively, the data support the hypothesis that TLR-7 agonism can induce immune-mediated clearance of infected cells and is a potential therapeutic approach for control or elimination of HBV infection with therapy of finite duration.
In this study, only 3 HBV chronically infected animals were available for the project; all were chronically infected for more than 2 decades. This limited the scope of the study to proof-of-concept that TLR-7 stimulation could impact HBV chronic infection. We were not able to evaluate regimen optimization or combination with direct antiviral therapy or to determine whether extended dosing at the low dose or combination with antiviral agents could result in the same reduction in viral DNA and antigen-positive hepatocytes without an increase in liver enzymes. In this context, it is also important to note that the results of this study presented evidence for a "presystemic" TLR-7 response: low doses of GS-9620 were capable of inducing an antiviral immune response at the level of the liver, PBMC, and possibly gut-associated lymphoid tissue without induction of systemic IFN-α or its side effects. At the low dose, in both uninfected and infected animals, GS-9620 induced ISGs in the liver and/or in PBMC with no detectable increase in serum levels of IFN-α. Ongoing clinical trials are evaluating whether administration of GS-9620 at presystemic dose levels provides similar antiviral benefits alone, as recognized in this study, or in combination with direct-acting antivirals, without the side effects often associated with systemic administration of IFN-α. Identification of a well-tolerated, finite therapeutic regimen, including the possible combination of immune modulators, such as GS-9620, with direct-acting antiviral agents for the treatment and cure of chronic HBV infection in a significant and clinically relevant percentage of treated patients would be transformative for this disease.
|
|
| |
| |
|
|
|