| |
Persistence of Virus Reservoirs in ART-Treated SHIV-Infected Rhesus Macaques after Autologous Hematopoietic Stem Cell Transplant
|
| |
| |
Download the PDF here
Published: September 25, 2014
http://www.plospathogens.org/article/info:doi/10.1371/journal.ppat.1004406
"......The key findings of this study are the following: (i) autologous HSCT using apheresis products collected prior to infection is feasible in SHIV-infected RMs; (ii) as expected, the myeloablative TBI used for conditioning induced a massive reset of the lympho-hematopoietic compartment, consequently resulting in the depletion of 94.2-99.2% of circulating CD4+ T-cells; (iii) animals receiving autologous HSCT under ART exhibited a prompt and pronounced decline in the peripheral blood viral reservoir (with undetectable SHIV-DNA in PBMCs in two out of three RMs) and maintained undetectable SHIV-RNA viremia with the exception of a few minor blips; (iv) two of the three transplanted RMs showed a very rapid rebound of viremia after ART interruption; and (v) the third transplanted RM, who was sacrificed for clinical reasons at day fourteen post ART interruption, had no detectable virus in plasma, PBMCs, tonsils, and GI tract, low but detectable levels of SHIV-DNA in sorted peripheral CD4+ T-cells and lymph nodes, and moderate levels of SHIV-DNA in the spleen.....
Effect of ART interruption on virus replication
ART was interrupted after stem cell engraftment (between 78 and 128 days post-initiation, Table 1). As expected, a rapid viral rebound was observed in the plasma of the three control animals as early as one week post ART cessation (Figure 5A). Two out of the three transplanted animals also experienced a rapid plasma viral rebound post ART interruption. The remaining transplanted animal (T2) maintained an undetectable plasma viral load at two weeks post ART interruption (Figure 5A). Unfortunately, further time-points were not analyzed in this animal as he was euthanized due to progressive renal failure. As shown in Figure 5B, ART interruption led to an increase of the SHIV-DNA levels in the PBMCs of the two transplanted RMs who also experienced a plasma viral rebound. This rebound in PBMC SHIV-DNA was observed at the first assessment post-ART interruption in both animals (day 28 for T1 and day 15 for T3). Of note, no SHIV-DNA was detected in the PBMCs of RM T2 who also maintained undetectable plasma viral load at two weeks after ART interruption. However, further analyses of this animal revealed low but detectable levels of SHIV-DNA in sorted peripheral CD4+ T-cells obtained at the same time-point (i.e., two weeks after ART interruption at necropsy) (Figure 5C).
SHIV-DNA viral load in tissues after ART interruption
Several tissues were collected at necropsy including ileum, jejunum, colon, rectum, superficial and mesenteric lymph nodes as well as tonsils. SHIV-DNA levels in cell suspensions obtained from these tissues were quantified by PCR. As shown in Figure 6, low levels of SHIV-DNA were detected in the spleen and lymph nodes of the transplanted RM who maintained an undetectable peripheral viral load post ART interruption (T2) but not in the tonsils or gut compartments. Of note, we were able to detect SHIV-DNA in the gut and tonsils of the other two transplanted RMs (T1 and T3) who exhibited a rapid rebound of viremia after ART interruption.
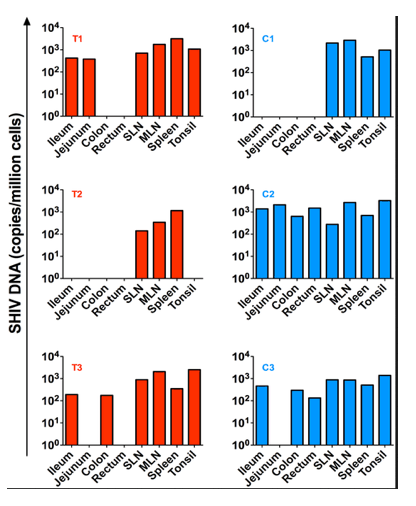
Figure 6. SHIV-DNA in tissues post ART interruption.
SHIV-DNA levels expressed as copies/million cells obtained at necropsy from the ileum, jejunum, colon, rectum, superficial and mesenteric lymph nodes, and tonsils are shown for each individual animal. Transplanted animals are depicted in red, controls in blue. Estimated number of CD4+ T-cells per million cells in tissues at necropsy is indicated in Table S1."
Abstract
Despite many advances in AIDS research, a cure for HIV infection remains elusive. Here, we performed autologous hematopoietic stem cell transplantation (HSCT) in three Simian/Human Immunodeficiency Virus (SHIV)-infected, antiretroviral therapy (ART)-treated rhesus macaques (RMs) using HSCs collected prior to infection and compared them to three SHIV-infected, ART-treated, untransplanted control animals to assess the effect of conditioning and autologous HSCT on viral persistence. As expected, ART drastically reduced virus replication, below 100 SHIV-RNA copies per ml of plasma in all animals. After several weeks on ART, experimental RMs received myeloablative total body irradiation (1080 cGy), which resulted in the depletion of 94-99% of circulating CD4+ T-cells, and low to undetectable SHIV-DNA levels in peripheral blood mononuclear cells. Following HSC infusion and successful engraftment, ART was interrupted (40-75 days post-transplant). Despite the observed dramatic reduction of the peripheral blood viral reservoir, rapid rebound of plasma viremia was observed in two out of three transplanted RMs. In the third transplanted animal, plasma SHIV-RNA and SHIV DNA in bulk PBMCs remained undetectable at week two post-ART interruption. No further time-points could be assessed as this animal was euthanized for clinical reasons; however, SHIV-DNA could be detected in this animal at necropsy in sorted circulating CD4+ T-cells, spleen and lymph nodes but not in the gastro-intestinal tract or tonsils. Furthermore, SIV DNA levels post-ART interruption were equivalent in several tissues in transplanted and control animals. While persistence of virus reservoir was observed despite myeloablation and HSCT in the setting of short term ART, this experiment demonstrates that autologous HSCT can be successfully performed in SIV-infected ART-treated RMs offering a new experimental in vivo platform to test innovative interventions aimed at curing HIV infection in humans.
Author Summary
While antiretroviral therapy (ART) can reduce HIV replication, it does not eradicate the virus from an infected individual. Replication-competent viruses persist on ART and our incomplete understanding of these viral reservoirs greatly complicates the generation of a cure for HIV. In this study we performed, for the first time, hematopoietic stem cell transplant (HSCT) in the established model of SIV infection of rhesus macaques (RM). The HSC originating from the bone marrow were collected before SIV infection. After SIV infection, RM were treated with ART for several weeks to reduce viral replication before performing a total body irradiation and a transplant with their own, pre-infection, stem cells. The irradiation eliminated 94-99% of the circulating CD4+ T-cells, the main cell target of HIV/SIV infection. A successful engraftment of the HSC was observed and blood viral reservoirs were drastically reduced. However, when ART was interrupted, a rapid rebound of plasma viremia was observed in two out of three transplanted RM indicating that the massive reset of the hematopoietic compartment was not sufficient to eliminate the total-body virus reservoir in the setting of short term ART. This model of HSCT in SIV-infected RM provides a new platform to investigate HIV eradication strategies.
Discussion
The apparent cure of HIV infection in the "Berlin patient" [5]-[7] has energized efforts to understand the mechanisms of virus persistence despite ART-mediated suppression of virus replication. The factors thought to be involved in the favorable outcome of the Berlin patient following HSCT include (i) the myeloablative conditioning regimen; (ii) the donor's homozygosity for ∼32ccr5; and (iii) the graft versus host effect. In this test-of-concept study of autologous HSCT in SHIV-infected RMs we interrogated the relative contribution of a myeloablative conditioning regimen in eliminating the persistent reservoir of latently infected cells. To the best of our knowledge this is the first time that a study of similar design has been conducted.
The key findings of this study are the following: (i) autologous HSCT using apheresis products collected prior to infection is feasible in SHIV-infected RMs; (ii) as expected, the myeloablative TBI used for conditioning induced a massive reset of the lympho-hematopoietic compartment, consequently resulting in the depletion of 94.2-99.2% of circulating CD4+ T-cells; (iii) animals receiving autologous HSCT under ART exhibited a prompt and pronounced decline in the peripheral blood viral reservoir (with undetectable SHIV-DNA in PBMCs in two out of three RMs) and maintained undetectable SHIV-RNA viremia with the exception of a few minor blips; (iv) two of the three transplanted RMs showed a very rapid rebound of viremia after ART interruption; and (v) the third transplanted RM, who was sacrificed for clinical reasons at day fourteen post ART interruption, had no detectable virus in plasma, PBMCs, tonsils, and GI tract, low but detectable levels of SHIV-DNA in sorted peripheral CD4+ T-cells and lymph nodes, and moderate levels of SHIV-DNA in the spleen.
Due to many logistical challenges of this experiment we chose to conduct the study in a temporally compressed fashion, with 37-53 days of ART before autologous HSCT, and interruption of ART after hematopoietic reconstitution, rather than prolonged continuation of therapy. This study was therefore designed to determine the impact of myeloablative irradiation on the viral reservoir, rather than the impact of prolonged viral suppression in conjunction with myeloablation. It is therefore possible that a similarly designed study, in which ART is maintained for a significantly longer period both before and after autologous HSCT, would have a different outcome, possibly demonstrating a more dramatic effect of autologous HSCT on the persistent reservoir of latently infected cells. Moreover, we cannot rule our the possibility that the level of virus suppression achieved by the short-term ART regimen in this experiment might not be as complete as what is observed in HIV-infected individuals on long-term ART. In this model of SHIV-infected RM, 5 to 7 weeks on ART pre-transplant may have been insufficient to fully suppress viral replication and the transient low-level viremia observed immediately post-transplant could be attributed to an insufficient period of ART pre-transplant. However, similar viral blips were observed in one patient who received allogeneic stem cell transplant after many years on combined ART [8]. Although the origin of these transient blips is unknown, it may represent release of the virus from latently infected cells in the setting of cell activation during conditioning and the peri-transplant period. In keeping with this hypothesis, it should be noted that in our study the post-transplant period was characterized by an expansion of CD4+ T-cells expressing CCR5 as well as proliferation and activation markers.
Together with the observed increased proportion of memory CD4+ T-cells post-transplant, these results suggest that the CD4+ T-cell compartment recovered primarily through homeostatic proliferation of memory CD4+ T-cells.
The myeloablative TBI used for conditioning resulted in the depletion of 94.2-99.2% of circulating CD4+ T-cells. Unfortunately, due to the clinical challenges of this innovative experiment, no tissue biopsies could be obtained immediately post-transplant to evaluate the TBI-induced CD4+ T-cell depletion in tissues. However, this study shows that myeloablative TBI and autologous HSCT did not prevent a rebound of viremia post-ART interruption in two out of three RMs despite relatively early ART initiation (day 28 post-infection). Moreover, while the SHIV-DNA level in PBMCs was undetectable or close to undetectable post autologous HSCT, it rapidly rebounded after ART interruption to levels that were similar or higher than those observed in the control animals at the same time-point. While in the third animal (T2) there was no sign of virus present in the plasma, PBMCs, and various tissues at the time of necropsy, this RM had to be sacrificed due to kidney failure at day fourteen after ART interruption making the interpretation of these data somewhat difficult. Of note, this study was not designed to identify the cellular and anatomic sources of the rapid plasma viral rebound observed in two transplanted RMs following ART interruption. Determining the relative contribution of tissue CD4+ T-cells, macrophages, and potentially other sources represents an important area for future investigation, amenable for interrogation with this model.
We acknowledge a number of limitations in our study including the small number of animals and the foreshortened time line involved. However, the demonstrated feasibility of this test-of-concept study in a non-human primate model of AIDS virus infection is per se an important result given the extreme complexity of the experimental protocol. The RMs included in this study underwent a series of procedures that have been only rarely, if ever, used in the same animal, including stem cell mobilization and harvesting by apheresis, RT-SHIV infection, daily four-drug ART administration, total body irradiation, re-infusion of HSCs, repeated platelet transfusions, and receipt of several antimicrobial prophylaxes. The feasibility of HSCT in SIV- or SHIV-infected RMs suggests, in our view, that further studies using this model in conjunction with longer term ART as well as additional interventions aimed at purging both the peripheral blood and lymphoid tissue-based viral reservoirs will provide critical information for the requirements to cure HIV infection in humans.
With respect to our understanding of the mechanisms responsible for "curing" HIV infection in the Berlin patient, our study supports the hypothesis that myeloablative TBI can cause a significant decrease in the viral reservoir in circulating PBMCs, even though it was not sufficient to eliminate all reservoirs. While the conditioning regimen in the Berlin patient also included antithymocyte globulin and chemotherapy, the use of a ∼32ccr5 homozygous donor and/or the presence of graft versus host disease likely played a significant role in that clinical context. The importance of graft versus host disease that effectively results in a "graft versus reservoir" effect is also emphasized by the recent observation of two HIV-infected patients in which a prolonged (i.e., 3-8 months) period of undetectable viremia in absence of ART was observed after allogeneic HSCT from donors with wild-type ccr5 alleles [9], although these patients did eventually develop rebound of viremia [16]. Future studies of allogeneic HSCT in SIV- or SHIV-infected RMs in the presence or absence of gene therapy interventions to knock out ccr5 would be very informative in this regard, and may elucidate the mechanism of the sustained cure seen in the Berlin patient but not the above mentioned recipients of donor cells wild type for ccr5.
In conclusion, we have conducted the first test-of-concept study of myeloablative irradiation and autologous HSCT in ART-treated SHIV-infected RMs. This experiment demonstrated that autologous HSCT is a feasible intervention that can lead to a marked reduction of the virus reservoir in the peripheral blood, and can be used as an experimental in vivo platform to test innovative interventions aimed at curing HIV infection in humans.
Introduction
The introduction of antiretroviral therapy (ART) has dramatically reduced the morbidity and mortality associated with HIV infection and AIDS. However, currently available ART requires life long treatment with significant potential side effects and a cost that places an inordinate burden on public health systems. While reduction of HIV viral loads below detectable limits is often achieved in ART-treated individuals, a treatment that can eradicate or functionally cure HIV infection remains elusive. Many studies indicate that the key obstacle to cure HIV infection is the presence of a persistent reservoir of latently infected cells that are not eliminated by ART [1], [2]. Thus, interruption of ART consistently results in a rebound of viremia to pre-treatment levels [3], [4]. Several biological aspects of this virus reservoir, including its exact cellular and anatomic origin as well as the mechanisms responsible for its establishment and persistence under ART remain poorly understood. This limited knowledge represents a fundamental barrier to a cure for HIV infection, and novel therapeutic strategies aimed at eliminating the reservoir will likely not be developed until we overcome this barrier.
In 2009 it was reported that an HIV-infected individual with acute myelogenous leukemia treated with myeloablative chemotherapy and allogeneic hematopoietic stem cell transplant (HSCT) from a ∼32ccr5 homozygous donor had remained without detectable HIV replication in the absence of ART for 1.8 years [5], [6]. This first demonstration of a functional cure in this patient was confirmed in 2013 in a follow-up study showing no signs of recrudescent HIV replication and waning of HIV-specific immune responses five years after interruption of ART [7]. More recently, two HIV-infected individuals have been described with prolonged (i.e., 3-8 months) suppression of viremia in absence of ART following allogeneic HSCT from donors homozygous for the wild type ccr5 allele [8], [9]. Similar to the "Berlin patient" described above, these two transplant recipients were themselves ∼32ccr5 heterozygotes. The factors involved in the lack of detectable virus replication after ART interruption in HIV-infected individuals undergoing HSCT are complex, and may include (i) the myeloablative regimen involving various combinations of chemotherapy, immunosuppression, and total body irradiation (TBI); (ii) the deficiency of CCR5 in the transplanted donor cells (in the first case); and (iii) a graft versus host effect that may target cells that are latently infected with HIV (i.e., graft versus reservoir effect). Assessing the relative contribution of these factors will likely provide useful information to define the clinical potential of HSCT as a cure for HIV infection.
SIV infection of non-human primates, such as rhesus macaques (RMs) has been used for over two decades as an in vivo model for studies of HIV pathogenesis, prevention, and treatment [10]. SIV-infected RMs show remarkable similarities to HIV-infected individuals in terms of mechanisms and markers of disease progression, and current ART regimens can fully suppress virus replication in these animals [11]-[14], thus making this model suitable for probing HIV eradication strategies. In this study, we conducted a controlled test of the contribution of pre-transplant myeloablative irradiation to clearance of the viral reservoir in a cohort of RMs infected with a chimeric simian-human immunodeficiency virus (SHIV) and treated with ART. To the best of our knowledge, this is the first time HSCT has been utilized in RMs to investigate viral persistence. The procedure was successfully performed after SHIV infection and ART-induced control of virus replication using HSCs collected prior to infection. While these recipients showed undetectable plasma viremia and low to absent SHIV-DNA in PBMCs after HSCT, interruption of ART resulted in a rapid rebound of virus replication in two out of three animals. The one transplanted RM who maintained undetectable viremia and SHIV-DNA PCR in PBMCs after ART interruption showed low but detectable levels of SHIV-DNA in sorted circulating CD4+ T-cells, spleen and lymph nodes but not in the gastro-intestinal tract or tonsils. Collectively, these results indicate that the massive reset of the lympho-hematopoietic compartment that follows TBI-induced myeloablation was not sufficient to eliminate the total-body virus reservoir in SHIV-infected RMs in the setting of short term ART. However, this study provides a critical foundation upon which to test other potential contributors to a transplant-mediated cure of HIV.
Results
Experimental design
Six RMs were included in this study. All six RMs were males with an average age of 4.2 years (Table 1). Figure 1 shows an overview of the experimental design. Three rhesus macaques (T1, T2, T3) were treated with G-CSF for CD34+ stem cell mobilization followed by HSC collection by leukopheresis and cryopreservation of the collected cells. The six RMs were infected i.v with 104 TCID50 RT-SHIVTC. Starting at week four post-infection all six RMs were initiated on ART. The ART regimen consisted of two nucleotide/side reverse transcriptase inhibitors (PMPA/tenofovir and FTC/emtricitabine), one non-nucleoside reverse transcriptase inhibitor (efavirenz) and one integrase inhibitor (raltegravir). After five to eight weeks on ART, RMs T1-T3 received myeloablative TBI as pre-transplant conditioning. The leukopheresis products collected before infection were infused within 24 hours following the last dose of TBI. Recipients were given a total of 7.3x108+/-1.3x108 total nucleated cells (TNC)/kg which corresponded to 2.9x106+/-1.1x106 CD34+ cells/kg. After successful engraftment of donor cells (five to eleven weeks post-transplant), ART was interrupted in RMs T1-T3 as well as in the control RMs.
ART control of RT-SHIV replication
As shown in Figure 2A, following experimental infection with RT-SHIVTC the six RMs experienced a rapid, exponential increase in virus replication that peaked at week two post infection (105-107 SHIV-RNA copies/ml plasma). ART initiated at week four after infection drastically reduced plasma viral load to less than 100 copies of SHIV-RNA per ml of plasma. Consistent with prior studies of SIV/SHIV infection in RMs, the absolute number of peripheral CD4+ T-cells was decreased following infection and partially restored on ART (Figure 2B).
Autologous HSCT: Conditioning and engraftment
The myeloablative TBI resulted in a drastic reduction of the absolute count of blood cells including neutrophils, monocytes, lymphocytes and CD4+ T-cells (Figure 3A). The nadir was observed at day eleven post-TBI for neutrophils (41.6-78.2 neutrophils/μl), day seven post-TBI for monocytes (4.4-14.8 monocytes/μl), and between day one and day five post-TBI for lymphocytes and CD4+ T-cells (54-60 lymphocytes/μl and 6.7-45.5 CD4+ T-cells/μl). Of note, between 94.2 and 99.2% of circulating CD4+ T-cells were eliminated by the TBI (Figure 3A). Engraftment was demonstrated by increasing neutrophil and platelet counts unsupported by transfusion. Neutrophil engraftment was defined as an absolute neutrophil count (ANC) exceeding 500 cells/μl for three consecutive days. The first of these three consecutive days was considered the day of engraftment. As shown in Figure 3B, neutrophil engraftment was successfully achieved between day sixteen and day eighteen post-HSC infusion in the three transplanted animals. During HSCT, the three transplanted animals received platelet and whole blood transfusions for thrombocytopenia prior to platelet engraftment, as well as several antimicrobial prophylactic interventions (Figure S1). Platelet engraftment was defined as a blood platelet count exceeding 20,000 cells/μl in absence of transfusion support for seven consecutive days. According to this definition, platelet recovery was achieved at 42, 22 and 33 days post-transplant for T1, T2 and T3, respectively (Figure 3C).
Effect of autologous HSCT on CD4+ T-cells
Following transplantation and engraftment, we observed a rapid increase in the absolute leukocyte count and a slower reconstitution of the circulating CD4+ T-cells (Figure 4A and B). The peripheral reconstitution of CD4+ T-cells appeared to involve peripheral T-cell expansion as evidenced by the increased proportion of circulating CD4+ T-cells expressing the proliferation antigen Ki-67 (Figure S2A). In addition, HLA-DR and CCR5 were increased on CD4+ T-cells following HSCT (Figure S2B,C). Further immunophenotypic analyses revealed a significant increase in the proportion of memory CD4+ T-cells (including memory stem cells, central memory, and effector memory) following transplantation (p = 0.03, Figure S3), similar to previous reports of both autologous and allogeneic HSCT [6], [15]. These results are consistent with CD4+ T-cells recovery occurring primarily through the homeostatic proliferation of memory CD4+ T-cells post-transplant.
Effect of myeloablative irradiation-based pre-transplant conditioning on virus replication and the peripheral blood viral reservoir
A few blips of transient low-level viremia were observed in the plasma of the three transplanted animals immediately after TBI and HSC infusion and while still on ART (Figure 5A). The origin of these transient increases in viral load is not clear, but it may represent release of virus from pre-existing reservoirs in the setting of events of CD4+ T-cell activation during conditioning and the peri-transplant period. With the exception of these transient episodes of viremia, the plasma viral load remained undetectable in all six animals on ART (Figure 5A). Of note, the ART regimen alone reduced the level of SHIV-DNA in PBMCs (i.e., the peripheral viral reservoir) by 1.0-1.5 log in the three control RMs (Figure 5B). In the transplanted animals, the reduction in cell-associated viral DNA was more pronounced, with two RMs showing levels of SHIV-DNA in PBMCs below the limit of detection and one RM (T1) close to this level (as low as 130 copies/million PBMC, Figure 5B). The normalization of the cell-associated SHIV-DNA level to the CD4+ T-cells counts suggest a decrease in the frequency of infection of these cells post-transplant (Figure 5B).
Effect of ART interruption on virus replication
ART was interrupted after stem cell engraftment (between 78 and 128 days post-initiation, Table 1). As expected, a rapid viral rebound was observed in the plasma of the three control animals as early as one week post ART cessation (Figure 5A). Two out of the three transplanted animals also experienced a rapid plasma viral rebound post ART interruption. The remaining transplanted animal (T2) maintained an undetectable plasma viral load at two weeks post ART interruption (Figure 5A). Unfortunately, further time-points were not analyzed in this animal as he was euthanized due to progressive renal failure. As shown in Figure 5B, ART interruption led to an increase of the SHIV-DNA levels in the PBMCs of the two transplanted RMs who also experienced a plasma viral rebound. This rebound in PBMC SHIV-DNA was observed at the first assessment post-ART interruption in both animals (day 28 for T1 and day 15 for T3). Of note, no SHIV-DNA was detected in the PBMCs of RM T2 who also maintained undetectable plasma viral load at two weeks after ART interruption. However, further analyses of this animal revealed low but detectable levels of SHIV-DNA in sorted peripheral CD4+ T-cells obtained at the same time-point (i.e., two weeks after ART interruption at necropsy) (Figure 5C).
SHIV-DNA viral load in tissues after ART interruption
Several tissues were collected at necropsy including ileum, jejunum, colon, rectum, superficial and mesenteric lymph nodes as well as tonsils. SHIV-DNA levels in cell suspensions obtained from these tissues were quantified by PCR. As shown in Figure 6, low levels of SHIV-DNA were detected in the spleen and lymph nodes of the transplanted RM who maintained an undetectable peripheral viral load post ART interruption (T2) but not in the tonsils or gut compartments. Of note, we were able to detect SHIV-DNA in the gut and tonsils of the other two transplanted RMs (T1 and T3) who exhibited a rapid rebound of viremia after ART interruption.
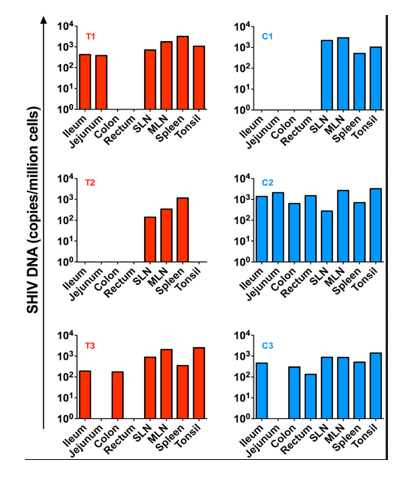
Figure 6. SHIV-DNA in tissues post ART interruption.
SHIV-DNA levels expressed as copies/million cells obtained at necropsy from the ileum, jejunum, colon, rectum, superficial and mesenteric lymph nodes, and tonsils are shown for each individual animal. Transplanted animals are depicted in red, controls in blue. Estimated number of CD4+ T-cells per million cells in tissues at necropsy is indicated in Table S1.
|
|
| |
| |
|
|
|