 |
|
|
| |
Defining total-body AIDS-virus burden with implications for curative strategies
|
| |
| |
Download the PDF here
Age, Inflammation, Blood Brain Barrier Permeability and Single Nucleotide Polymorphisms in Transporters May Influence Cerebrospinal Fluid Antiretrovirals' Concentrations
Jacob D Estes1, Cissy Kityo2, Francis Ssali2, Louise Swainson3, Krystelle Nganou Makamdop4, Gregory Q Del Prete1,
Steven G Deeks5, Paul A Luciw6, Jeffrey G Chipman7, Gregory J Beilman7 , Torfi Hoskuldsson7,
Alexander Khoruts8, Jodi Anderson8, Claire Deleage1, Jacob Jasurda8, Thomas E Schmidt8, Michael Hafertepe8,
Samuel P Callisto8 , Hope Pearson8, Thomas Reimann8, Jared Schuster8, Jordan Schoephoerster8,
Peter Southern9, Katherine Perkey9, Liang Shang9, Stephen W Wietgrefe9, Courtney V Fletcher10,
Jeffrey D Lifson1, Daniel C Douek4, Joseph M McCune3, Ashley T Haase9 & Timothy W Schacker8
The density of vRNA+ cells was highest in LT, but vRNA+ cells were detected in every organ, including the heart, liver, and kidney, and the central nervous system. To estimate the total virus burden for different tissues, we assumed that each organ contributes vRNA+ cells to the total population of vRNA+ cells in an amount proportional to the frequency of vRNA+ cells measured by ISH and the total mass of these organs. In the RMs studied at peak viremia, for example, the mass of the heart was approximately 38.3 g, and the frequency of vRNA+ cells was 7.54 X 103 cells/g; thus, the total number of infected cells in this organ was accordingly estimated at ∼2.89 X 105 cells (Supplementary Table 1). The combined mass of lymph nodes for this animal was estimated to be 78 g (∼1% of total body mass). With a mean frequency of ∼8.73 X 105 vRNA+ cells/g, the total population of vRNA+ cells in LNs was estimated at 7.68 X 107 cells, approximately 2 log10 higher than that found in the heart. In the gut, we measured ∼5.57 X 105 vRNA+ cells/g; thus, on the basis of the total organ weight of 315 g in this animal, ∼1.68 X 108 vRNA+ cells, or more than two-thirds of the total number of vRNA+ cells in all of the organs examined, were in the gut. The contribution of each organ to the estimated total population of infected cells (as a percentage of the total) before and during ART (graphically shown in Fig. 1) illustrated that the primary reservoir sites of infection were LTs, in which ∼98.4% of the vRNA+ cells resided (gut, LNs, and spleen). The addition of the lung, a tissue with abundant mucosal-associated LT, increased our estimate of the frequency of vRNA+ cells residing in LTs to ∼99.6%.
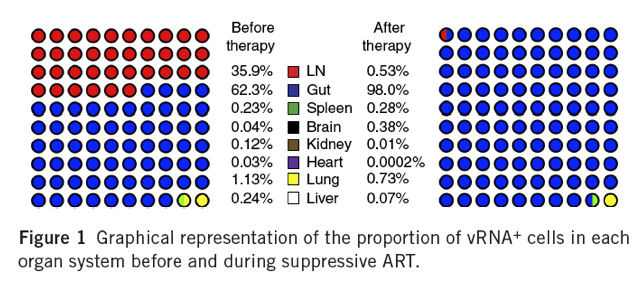
Effects of ART on vRNA+ and vDNA+ cell populations in tissues
In SIVmac251-infected RMs that received 20-22 weeks of ART beginning at 56 dpi, as compared with those with untreated infections, the frequency of vRNA+ cells decreased by approximately 2 log10 in LNs (Fig. 3a and Supplementary Table 2a-d) but by only approximately twofold or less in the gut and spleen. The frequency of vRNA+ cells was actually higher in the brains of animals receiving ART compared with untreated animals, a result presumably reflecting interanimal variability (Supplementary Table 2a) and poor penetration of drugs into the brain. In the RT-SHIV-infected animals, 26 weeks of ART decreased the number of vRNA+ cells by ∼3 to 4 log10 in all LT, but by only approximately twofold in the brain (Fig. 3b). The greater decreases seen in the RT-SHIV-infected animals compared with SIV-infected animals may reflect the relative effectiveness of the different ART regimens against the respective target viruses, distinct differences in viral replication capacity in RMs, differences in tissue concentrations of ART, or Env-specific immune responses.
We have previously shown in HIV-infected humans that the intracellular concentrations of antiretroviral drugs (ARVs) in LT can be significantly less than the concentrations in peripheral blood mononuclear cells (PBMCs) and well below the optimal concentration for viral suppression5. We measured intracellular concentrations of ARVs in PBMCs and cells of the LN, ileum (gut-associated lymphoid tissue (GALT)) and rectal-associated mucosal tissue (RALT) from six of the animals described above and similarly found lower ART concentrations in LT than in blood (Fig. 4). The median ratios for TFV-diphosphate (TFV-DP, an active phosphorylated form of the drug) were 0.84 for LN/PBMCs, 0.41 for GALT/PBMCs, and 0.84 for RALT/PBMCs. The median ratios for FTC-triphosphate (FTC-TP) were 0.26 for LN/PBMCs, 0.32 for GALT/PBMCs, and 0.32 for RALT/PBMs. The median RALT/PBMC ratio for DRV was 15; however, DRV was not quantifiable in LN or GALT samples. Thus, the concentrations of the ARVs were not equivalent to those in peripheral blood in any of the studied compartments, with the exception of DRV in RALT (summary of intracellular concentration of ARVs for all drugs studied in Supplementary Table 3). These results support the hypothesis that comparatively lower ARV concentrations in different tissues may contribute to incomplete suppression of viral replication in those tissues.
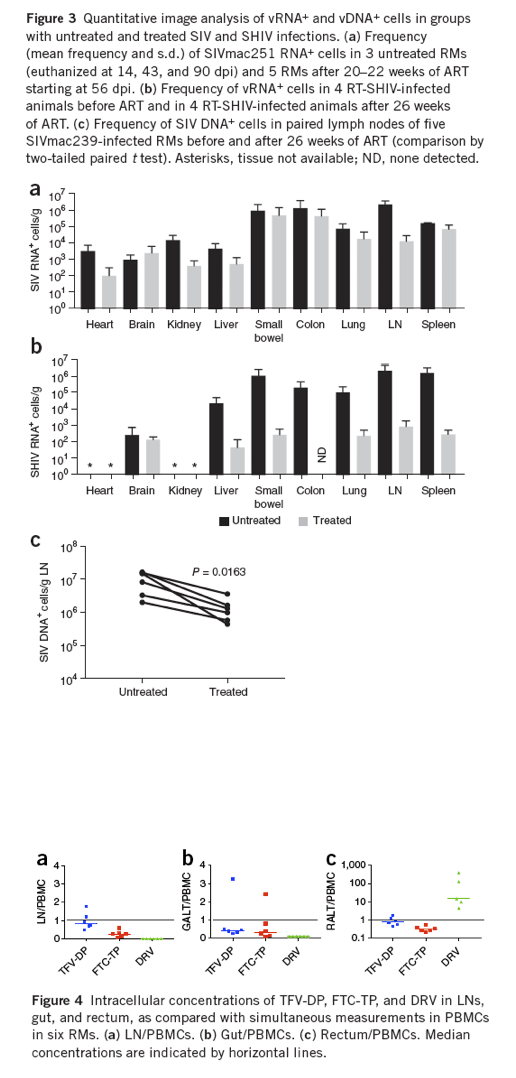
With the exception of the heart tissues, vRNA+ cells were detectable in every animal on ART, even though all had plasma viral loads <50 copies/mL (the limit of detection of the assay used). The highest numbers of vRNA+ cells during ART were mainly in LTs (LN, spleen, and GALT) and the lung. Because detection of vRNA+ cells signifies transcription and not necessarily virus production, we used ISH/TSA amplification with an enzyme-linked immunofluorescence technique (ELF97) to detect virus-producing cells through light microscopy23, to show that vRNA+ cells also produced virus during ART. We measured the frequency of vRNA+ cells with RNAscope and the frequency of cells producing virions with TSA/ELF97 in 20 subjacent sections from two of the SIVmac251-infected animals on suppressive ART for >6 months (Fig. 5). There was good agreement between the two measurements (Supplementary Table 4), a result consistent with the conclusion that most vRNA+ cells detected in LN were producing virions.
On the basis of preliminary analyses in which we found that clade B riboprobes detected fewer vRNA+ cells than did riboprobes for the prevalent clades A and D in Uganda (data not shown), we designed RNAscope and DNAscope probes specific to clades A and D for this study. With these probes, we determined an average frequency of vRNA+ cells in the LN and rectum before ART of ∼9.4 X 104 cells/g (6.3 X 103 to 3.5 X 105 cells/g) and ∼2.5 X 105 cells/g (2.3 X 104 to 9.6 X 105 cells/g), respectively (Fig. 6a), a result in reasonable agreement with previous estimates of ∼5 X 104/g in LNs of HIV-infected patients in the United States, with a mean CD4+ T cell count of 400 cells/μl (ref. 16). However, this value was more than tenfold lower than that observed in the SIVmac251-infected RMs necropsied at 90 dpi.
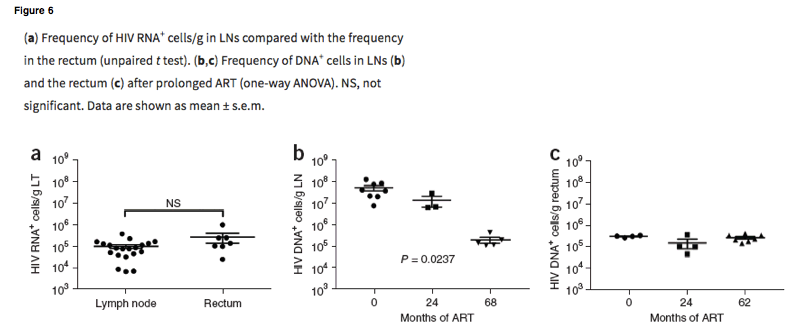
ART had a significant effect on the LT reservoir in individuals who had been treated for >2 years (range 2-6 years) and had undetectable plasma viral loads (<40 copies/mL in the first 12 months of the study and then <20 copies/mL). The mean frequency of vDNA+ cells declined by ∼3 log, to ∼2.7 X 105 cells/g (range 1.2 X 105 to 4.5 X 105 cells/g; P = 0.0237, analysis of variance (ANOVA)). In gut tissues, however, we did not observe a significant decrease in the size of the vDNA+-cell reservoir (Fig. 6c): before ART, the frequency of vDNA+ cells was ∼3.2 X 105 cells/g LT (range 2.8 X 105 to 3.4 X 105) compared with a mean of ∼2.7 X 105 cells/g (range 1.5 X 105 to 4.3 X 105 cells/g) after >62 months of ART (range 2-6 years; P > 0.05).
Abstract
In the quest for a functional cure or the eradication of HIV infection, it is necessary to know the sizes of the reservoirs from which infection rebounds after treatment interruption. Thus, we quantified SIV and HIV tissue burdens in tissues of infected nonhuman primates and lymphoid tissue (LT) biopsies from infected humans. Before antiretroviral therapy (ART), LTs contained >98% of the SIV RNA+ and DNA+ cells. With ART, the numbers of virus (v) RNA+ cells substantially decreased but remained detectable, and their persistence was associated with relatively lower drug concentrations in LT than in peripheral blood. Prolonged ART also decreased the levels of SIV- and HIV-DNA+ cells, but the estimated size of the residual tissue burden of 108 vDNA+ cells potentially containing replication-competent proviruses, along with evidence of continuing virus production in LT despite ART, indicated two important sources for rebound following treatment interruption. The large sizes of these tissue reservoirs underscore challenges in developing 'HIV cure' strategies targeting multiple sources of virus production.
What are the cellular and anatomic compartments from which infection might rebound after ART and achieve a functional cure or eradication? To answer that question, we performed an extensive analysis of potential sources of viral rebound, using both established and next-generation in situ hybridization (ISH) technologies and quantitative image analysis. We paid particular attention to LTs in HIV and simian immunodeficiency virus (SIV) infections, in which CD4+ T cells in the gut and secondary LT have previously been shown to be the principal sites where most of the cells producing viral (v) RNA (vRNA+ cells) are found in HIV-infected humans and SIV-infected macaques2,3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18, and where there is a very large pool (more than 1010 cells) of transcriptionally silent vDNA+ cells in LTs3, some of which contain replication-competent proviruses.
We analyzed brain, gut, heart, kidney, liver, lung, LN, and spleen tissues collected in necropsies of eight RMs infected with SIVmac251 (three untreated and five subjected to ART) and housed at Advanced Bioscience Laboratories.
We quantitatively characterized cellular tissue reservoirs in untreated SIV infection in the brain, heart, kidney, liver, lung, spleen, LNs (axillary, colonic, inguinal, and mesenteric), and multiple sites in the gut (duodenum, jejunum, ileum, ascending and transverse colon, and rectum) (representative images of SIV in situ hybridization from multiple tissues through two complementary approaches in Supplementary Fig. 3).
|
| |
|
|
|
|
|