| |
Inflammation Marker D-Dimer Associated with cell-associated HIV RNA in Virally Suppressed HIV+..... Aging is associated with inflammation and non-significantly higher ca-HIV RNA levels
|
| |
| |
Download the PDF here
"While current antiretroviral treatments for HIV are highly effective, data has shown that people living with HIV appear to experience accelerated aging and have shorter lifespans -- by up to five to 10 years -- compared to people without HIV. These outcomes have been associated with chronic inflammation, which could lead to the earlier onset of age-associated diseases, such as atherosclerosis, cancers, or neurocognitive decline.....Even with suppressive cART, some but not all studies suggest PWH have shorter life expectancy as compared to age-matched uninfected individuals [7, 8]. The increased mortality among PWH has been attributed to a higher incidence of age associated morbidities that potentially emerge as a result of chronic inflammation .....Aging is associated with inflammation and non-significantly higher ca-HIV RNA levels....Together these data suggest that aging is correlated with lower HIV-1 transcriptional control, although non-virus processes may also be impacting the associated higher inflammation observed in older PWH.....Collectively, our observations suggest that enhanced inflammation observed with aging does not promote greater HIV-1 transcription, but rather the presence of HIV-1 RNA in the absence of any new rounds of infection promotes innate immune activation .....Our model argues that novel drugs that lower the levels of intracellular HIV-1 RNA, something that current antiretrovirals cannot accomplish, may impact the chronic inflammation observed in PWH, especially as they age. ".... To our knowledge, no prior study has examined associations with intact proviral DNA levels and markers of inflammation. Intact proviruses may potentially induce greater host immune responses because of low level spreading infection or higher expression of structurally-intact virus proteins. .....The multivariable linear regression analysis also suggested that with every year of age, there was 0.02 log10 increase in plasma D-dimer level (Table 2). The median of all six measured plasma inflammatory markers was higher in the older as compared to the younger group (Table 3), although after correction for multiple comparisons, only the D-Dimer (unadjusted p = 0.0006, BH p-value 0.004, Fig. 2A) difference was statistically significant. .......ca-RNA was higher in the older as compared to younger group although the difference only showed a statistical trend (p = 0.08, Fig. 2B). ....In multivariable linear regression analysis, one year of older age was associated with 0.007 log10 higher ca-HIV-1 RNA (p = 0.11) after adjusting for duration of cART therapy .....although not statistically significant, ca-RNA trended higher in the older as compared to younger PWH. .....The directionality of the association between systemic inflammation and HIV transcription remains uncertain. Although not mutually exclusive, an inflammatory milieu may drive HIV-1 transcription or alternatively intracellular HIV-1 RNA expression may induce inflammation. ......The level of HIV-1 transcription was also measured in the cell lines in the presence and absence of TNF-alpha because this cytokine has been previously shown to stimulate HIV-1 RNA transcription [27, 28]. HIV-1 RNA was detectable in both cell lines over a range of TNF-alpha concentrations demonstrating that HIV-1 transcription could be induced with minimal cytokine stimulation. (Supplementary Figure 1A and 1B). .....This suggests that the higher pre-existing inflammation present in plasma does not induce greater HIV-1 transcription in these two different cell lines.
• despite the effectiveness of cART in preventing viremia, low level expression of cell-associated (ca) HIV-1 RNA remains present
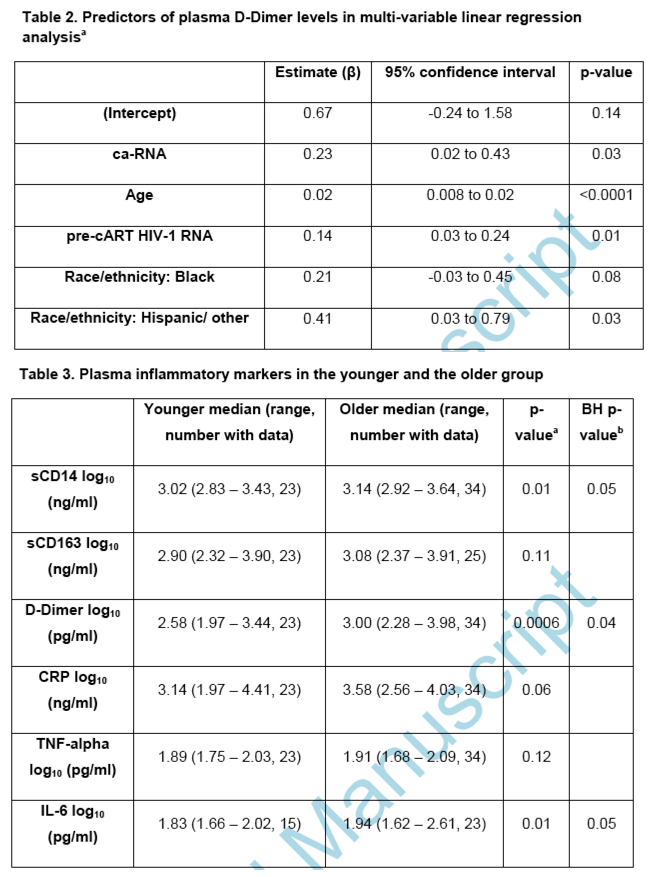
• In this work, we evaluated the association between age, chronic inflammation, and levels of intact proviral DNA and total ca-RNA. We also examined if plasma with varying levels of inflammation can differentially induce HIV-1 RNA expression in latently infected cells. ....We examined 57 HIV-1 cART treated and virologically suppressed individuals who were between the ages of 18-35 (n = 23) or ≥50 (n = 34) years (Table 1)....Plasma D-dimer levels showed a significant correlation with ca-HIV-1 RNA levels (ρ= 0.36, p = 0.006, Fig. 1A). Multivariable linear regression analysis was used to account for the baseline demographic differences among the older and younger group (Table 1). In the final model, ca-RNA level, age, pre-therapy log10 plasma HIV-1 copies, and ethnicity/race were predictive of plasma D-dimer levels (Table 2). Specifically, for every 10 fold increase in ca-RNA, there was a 0.23 log10 increase in plasma D-dimer level.
• PWH who have suppressed plasma viremia experience chronic systemic inflammation as compared to age-matched HIV-1 uninfected individuals.....To date, however, no study has examined the association between markers of inflammation and the number of intact, presumably infectious, HIV-1 proviruses. In PWH with sustained virus suppression, the majority of infected cells contain defective proviruses with deletions or hyper mutations [19, 20]. The intact and defective proviruses can yield virus RNA and proteins [21, 22], which potentially allows the host to recognize the provirus harboring cell. This immune response may account for a possible association between chronic inflammation and intact infectious provirus levels.
• Among HIV-1 negative individuals, there is a large body of literature supporting the notion that chronic systemic inflammation increases the risk for atherothrombosis, cancer, and other diseases associated with aging [5, 6]. In the current cART era, HIV-associated non-AIDS diseases, such as atherothrombosis, neurocognitive decline, and cancer, account for most of the morbidity and mortality in PWH who have suppressed virus levels [3]. Similar to HIV-1 uninfected individuals, these non-communicable HIV-associated non-AIDS diseases have been associated with elevated biomarkers, such as D-Dimer, C-reactive protein (CRP), soluble (s)-CD14, and interleukin (IL-6) [9-12].
• While current antiretroviral treatments for HIV are highly effective, data has shown that people living with HIV appear to experience accelerated aging and have shorter lifespans -- by up to five to 10 years -- compared to people without HIV. These outcomes have been associated with chronic inflammation, which could lead to the earlier onset of age-associated diseases, such as atherosclerosis, cancers, or neurocognitive decline.
• "Current antiretroviral drugs prevent new infection, but they do not prevent HIV RNA production, which our results point as a potential key factor driving inflammation in people living with HIV."
• Although combination antiretroviral therapy (cART) routinely reduces HIV-1 replication below levels detectable by conventional clinical assays, treatment does not eliminate infection. Infection persists in latently infected cells, and virus can re-emerge if cART is interrupted [1]. Further, despite the effectiveness of cART in preventing viremia, low level expression of cell-associated (ca) HIV-1 RNA remains present [2]. People with HIV (PWH) who are not on cART have high levels of systemic immune activation [3]. This inflammation, estimated via plasma cytokines / chemokine levels and cellular activation markers, decreases with cART-induced virus suppression, but it does not normalize to pre-infection levels [4]. Thus, PWH who have suppressed plasma viremia experience chronic systemic inflammation as compared to age-matched HIV-1 uninfected individuals.
• "We hope that our study results will serve as a springboard for examining drugs that stop HIV RNA production as a way to reduce inflammation,"
• "Our findings suggest that novel treatments are needed to target the inflammation persistent in people living with HIV,"
DISCUSSION
Even with suppressive cART, some but not all studies suggest PWH have shorter life expectancy as compared to age-matched uninfected individuals [7, 8]. The increased mortality among PWH has been attributed to a higher incidence of age associated morbidities that potentially emerge as a result of chronic inflammation [9-12]. The etiology of the chronic inflammation in individuals with suppressed virus level is likely multifactorial and specific drivers remain uncertain. In this study, we show that inflammation as determined by plasma markers associates with intracellular HIV-1 RNA but not intact proviral DNA levels. Furthermore, plasma inflammation increases with age in PWH, similar to what has been observed in HIV-1 uninfected individuals [33]. We also found that in-vitro HIV-1 RNA transcription in latent cell line models do not increase in the presence of plasma from older individuals who had greater levels of inflammatory markers. In aggregate, our observations suggest that as PWH age, they have increased systemic inflammation and potentially greater level of HIV-1 transcription, independent of intact proviral numbers. This age-associated inflammation, however, does not induce HIV-1 RNA transcription in latently infected cell lines. Aging is also not associated with higher levels of infectious virus. Our observations suggest that aging may be associated with greater uncontrolled HIV-1 RNA transcription, and this promotes chronic inflammation in the absence of new rounds of virus replication. Larger cohorts need to be examined to confirm our findings.
Previous studies have also examined the link between cellular or plasma markers of inflammation with HIV-1 intracellular DNA and RNA levels. These investigations have yielded conflicting data potentially due to cohort differences, such as the duration of cART and prospective versus cross sectional study design [13-18]. To our knowledge, no previous studies have examined intact proviral DNA, but instead have measured total or integrated DNA, and cell-associated RNA or plasma HIV-1 via ultrasensitive assays. Recently, it has been shown that intact proviral DNA, as opposed to ca-HIV-1 RNA and total HIV-1 DNA levels, correlates better with inducible infectious virus among virus suppressed PWH [20]. Similar intact provirus levels among younger as compared to older PWH on long-term cART implies that there is no difference in ongoing virus replication despite age. Thus, low level infectious virus production does not account for the enhanced plasma inflammation in older PWH.
In contrast to intact proviral DNA, we observed that ca-HIV-1 RNA, a measurement of transcriptional activity, showed a modest link with D-Dimer. The majority of the intracellular HIV-1 RNA as opposed to intact proviral DNA may constitute both infectious and non-infectious virus. First, majority of the persistent HIV-1 DNA in virus suppressed PWH is defective and will not yield infectious virus [19]. The integrated defective DNA, however, can still yield HIV-1 RNA, and in some cases this RNA is translated to viral proteins [21, 22]. Second, the small amount of HIV-1 RNA that may lead to infectious virions will likely not yield new productively infected cells because the presence of cART prevents spreading infection. It is important to note that current antiretroviral drugs prevent the generation of infectious virus and subsequent infection, but they do not inhibit HIV-1 transcription and RNA production. Thus, PWH on suppressive cART with no evidence of ongoing virus replication continue to have ongoing HIV-1 RNA production [2].
Recent studies suggest that the presence of HIV-1 RNA alone in the absence of protein or infectious virus induces inflammation. These recent investigations argue that nuclear export of HIV-1 intron containing RNA induces an innate immune response in CD4+ T cells, dendritic cells, and macrophages [34, 35]. In addition to these in-vitro HIV-1 studies, ex-vivo and animal studies also support the notion that endogenous retroviral RNA expression can induce inflammation [36]. This endogenous retroviral expression induced inflammation has been proposed as a contributor to age-associated inflammation, and aging has been associated with de-repression of endogenous retrovirus transcription [36]. Consistent with these data, we observed that HIV-1 RNA levels trended higher in the older as compared to the younger PWH even though the older group had been on suppressive cART for a longer time. This observed difference should not have a binary interpretation just because it was not statistically significant. Indeed, higher intracellular HIV-1 RNA levels have also been observed among suppressed HIV-1 older as compared to younger individuals in another study [14]. Although directionally consistent, a significant association between ca-HIV-1 RNA and D-dimer was only observed in the younger (ρ = 0.50, p = 0.01) but not the older PWH (ρ = 0.16, p = 0.37), possibly due to smaller sample sizes. Together these data suggest that aging is correlated with lower HIV-1 transcriptional control, although non-virus processes may also be impacting the associated higher inflammation observed in older PWH.
Our in-vitro studies further show that the greater inflammatory mediators present in plasma samples from older as compared to the younger PWH did not impact the HIV-1 transcriptional landscape in latent cells. Our observations would argue that pre-existing inflammatory milieu present in older PWH does not induce HIV-1 transcription in latently infected cells. We acknowledge that these in-vitro studies do not perfectly mimic the in-vivo conditions where majority of infected cells are present in tissues rather than peripheral blood and infected cell characteristics, such as integration site, impact transcriptional capacity. Collectively, our observations suggest that enhanced inflammation observed with aging does not promote greater HIV-1 transcription, but rather the presence of HIV-1 RNA in the absence of any new rounds of infection promotes innate immune activation [34, 35]. Our model argues that novel drugs that lower the levels of intracellular HIV-1 RNA, something that current antiretrovirals cannot accomplish, may impact the chronic inflammation observed in PWH, especially as they age.

The Journal of Infectious Diseases 19 October 2020
Alex Olson1, Carolyn Coote1, Jennifer E. Snyder-Cappione2, Nina Lin1,*, Manish Sagar1, *
1 Division of Infectious Diseases, Department of Medicine, Boston University School of Medicine, Boston, MA, USA
2 Department of Microbiology, Boston University School of Medicine, Boston, MA, USA
Abstract
HIV-1 infected individuals have increased inflammation, which has been associated with age-associated diseases. Plasma markers, cell-associated (ca) virus levels, and ability to stimulate RNA transcription in latently infected cell lines was examined in younger and older HIV-1 infected individuals with suppressed virus. ca-RNA, but not intact provirus level, had positive correlation with plasma D-Dimer levels. The older as compared to the younger group had higher D-dimer levels and a trend toward more ca-RNA, but similar levels of intact proviruses. Even though all measured inflammatory markers were relatively higher in the older as compared to younger individuals, this greater inflammation did not induce more HIV-1 transcription in latently infected cell lines. Inflammation and HIV-1 RNA expression increase with age despite similar levels of intact infectious HIV DNA. While plasma inflammation correlates with HIV-1 RNA expression in peripheral blood mononuclear cells, it does not induce HIV-1 transcription in latently infected cell lines.
RESULTS
Study Population
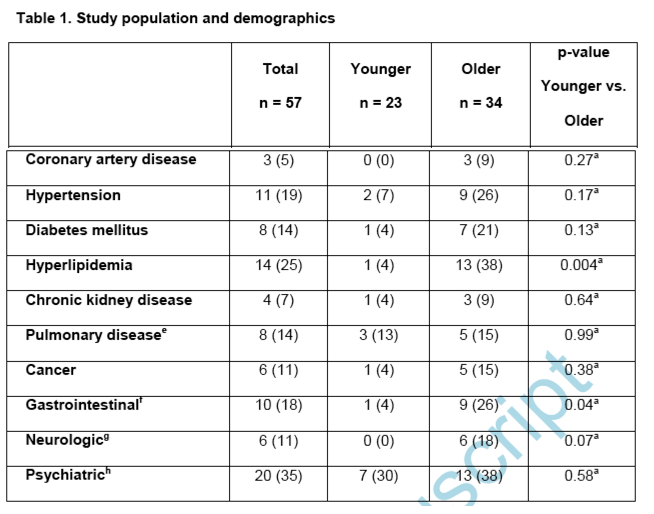
We examined 57 HIV-1 cART treated and virologically suppressed individuals who were between the ages of 18-35 (n = 23) or ≥50 (n = 34) years (Table 1). Women were part of the older but not the younger group. The younger group contained a significantly higher proportion of non-White, non-Black participants. The younger as compared to older group had higher pre-cART plasma HIV-1 levels and recorded nadir and enrollment absolute CD4 count although the differences were not statistically significant. The younger as compared to the older group had been on cART for a significantly shorter duration of time. The antiretroviral agent anchoring the cART regimen was not different among the groups. As expected, a greater number of the older as compared to younger individuals had pre-existing conditions.
Cell-associated HIV-1 RNA but not intact proviral DNA levels correlate with plasma inflammation.
We measured six different plasma inflammatory markers. Plasma D-dimer levels showed a significant correlation with ca-HIV-1 RNA levels (ρ= 0.36, p = 0.006, Fig. 1A). This D-dimer association remained statistically significant after adjusting for multiple comparisons (6 independent tests, BH p = 0.036 or for the Bonferroni method any unadjusted p-value < 0.008). Although there were no significant associations with the other plasma markers, there was a statistical trend with sCD14 (ρ= 0.23, p = 0.09) and sCD163 (ρ= 0.25, p = 0.09). sCD163 data, however, was not available from 9 participants.
Multivariable linear regression analysis was used to account for the baseline demographic differences among the older and younger group (Table 1). In the final model, ca-RNA level, age, pre-therapy log10 plasma HIV-1 copies, and ethnicity/race were predictive of plasma D-dimer levels (Table 2). Specifically, for every 10 fold increase in ca-RNA, there was a 0.23 log10 increase in plasma D-dimer level.
To our knowledge, no prior study has examined associations with intact proviral DNA levels and markers of inflammation. Intact proviruses may potentially induce greater host immune responses because of low level spreading infection or higher expression of structurally-intact virus proteins. We assayed intact proviral DNA in a median of 96,674 PBMCs (range 22,469 to 3,483,606). There was no significant difference in the number of cells examined among the older (median 87,124, range 22,469 to 412,421 cells) as compared to the younger group (median 107,645, range 29,568 to 3,483,606 cells, p = 0.23). Intact proviral DNA levels in bulk PBMC (median 2.1, range -0.01 – 3.4 log10 per 106 PBMC) were higher in our cohort than that previously documented in bulk PBMC from another cohort of virus suppressed PWH [30]. This observed variation could be attributed to differences in the duration of suppressive cART (median 3.3 versus 9 years) among the cohorts because intact proviruses may decay with prolonged cART [20]. There was a statistical trend in the association between the number of intact proviral DNA and sCD163 (ρ= 0.28, p = 0.05, Fig. 1B), but this was not adjusted for multiple comparisons. No associations were observed with the other plasma inflammatory markers.
Aging is associated with inflammation and non-significantly higher ca-HIV RNA levels
Higher plasma inflammatory markers has been observed with increasing age [31, 32]. The multivariable linear regression analysis also suggested that with every year of age, there was 0.02 log10 increase in plasma D-dimer level (Table 2). The median of all six measured plasma inflammatory markers was higher in the older as compared to the younger group (Table 3), although after correction for multiple comparisons, only the D-Dimer (unadjusted p = 0.0006, BH p-value 0.004, Fig. 2A) difference was statistically significant.
We hypothesized that ca-HIV-1 RNA, but not intact DNA, would be higher in the older as compared to the younger group because ca-RNA but not intact provirus level was associated with plasma inflammation (Fig. 1 and Table 2). Indeed, ca-RNA was higher in the older as compared to younger group although the difference only showed a statistical trend (p = 0.08, Fig. 2B). Older as compared to the younger PWH had longer duration of cART therapy (Table 1), and this prolonged drug exposure may impact ca-RNA levels. In multivariable linear regression analysis, one year of older age was associated with 0.007 log10 higher ca-HIV-1 RNA (p = 0.11) after adjusting for duration of cART therapy. The other baseline demographic characteristics did not have a significant association with ca-HIV-1 RNA. Thus although not statistically significant, ca-RNA trended higher in the older as compared to younger PWH.
On the other hand, intact proviral DNA level did not demonstrate a statistically trending difference among the two age-stratified groups (Fig. 2C). Age (β = .43, p = 0.67), duration of cART therapy (β = -0.31, p = 0.30) and absolute CD4 count at enrollment (β = -0.0001, p = 0.74) also did not predict the intact proviral DNA levels in univariate linear regression analyses. There were also no significant associations in multivariable linear regression analysis. Although not statistically significant, the negative correlation aligns with previous observation that intact provirus levels decrease with prolonged duration of cART-mediated virus suppression [20].
Plasma inflammation and aging does not enhance HIV-1 RNA production in latently infected cells
The directionality of the association between systemic inflammation and HIV transcription remains uncertain. Although not mutually exclusive, an inflammatory milieu may drive HIV-1 transcription or alternatively intracellular HIV-1 RNA expression may induce inflammation. Our observed association between ca-RNA level and plasma inflammation (Fig. 1 and Table 2) does not distinguish between these possibilities. We examined the impact of the plasma inflammatory milieu on HIV-1 transcription to further explore this relationship. We examined two different latently infected cell lines (J Lat 6.3 and ACH-2) because HIV-1 transcriptional capacity in latent cells varies depending on the integration site and other cell characteristics [30]. Thus, we expected varying results from the 2 different cell lines. Cell lines were incubated in media containing 10% plasma from PWH [27, 28]. A 10% plasma fraction was used because J-Lat 6.3 and ACH-2 cell cultures are routinely done with RPMI containing 10% FBS. The level of HIV-1 transcription was also measured in the cell lines in the presence and absence of TNF-alpha because this cytokine has been previously shown to stimulate HIV-1 RNA transcription [27, 28]. HIV-1 RNA was detectable in both cell lines over a range of TNF-alpha concentrations demonstrating that HIV-1 transcription could be induced with minimal cytokine stimulation. (Supplementary Figure 1A and 1B).
Although the older as compared to younger group had higher median levels of all the six measured inflammatory markers (Table 3), HIV-1 RNA relative to DNA levels trended higher among JLat cells incubated in plasma from the younger as compared to older PWH without (p = 0.05) but not with TNF-alpha (p = 0.23, Fig. 3A). RNA relative to DNA levels were also higher in ACH-2 cells incubated in plasma from the younger as compared to older PWH with (p = 0.04) but not without TNF-alpha (p = 0.38, Fig. 3B). As a positive control, higher HIV-1 RNA was observed in the presence as compared to the absence of TNF-alpha in both cell lines conditioned with participant plasma, implying that the patient plasma did not prevent HIV-1 transcription from the latent provirus (Fig. 3A and 3B). There was no correlation in the observed transcription induction in the presence of participant plasma among the two different cell lines (data not shown). This suggests that the higher pre-existing inflammation present in plasma does not induce greater HIV-1 transcription in these two different cell lines.
|
|
| |
| |
|
|
|