| |
Blunted muscle mitochondrial responses to exercise training in older adults with HIV
|
| |
| |
Download the PDF here
Blunted Increase in Muscle Mass After Exercise Training in People Aging with HIV
Do People With HIV Have to Exercise Harder to Get Same Muscle Benefit?
High-Intensity Exercise Has Edge Over Moderate Intensity in Older Group With HIV: Effects of HIV status on functional benefits of exercise in older sedentary adults. 25th Conference on Retroviruses and Opportunistic Infections (CROI). March 4-7, 2018
Antiretroviral initiation is associated with increased skeletal muscle area and fat content.....previous thinking that weight gain was return to normal is contradicted by this study saying the weight gain is fat.
Aging & Mitichondria decline
Summary: In older people with HIV skeletal muscle mitochondrial responses to moderate intensity exercise training were blunted compared to uninfected peers. Alternative exercise strategies, such as high-intensity interval training, may be necessary to further stimulate mitochondrial adaptations in people with HIV.
JID Dec 30 2020 - Catherine M. Jankowski1, Melissa P. Wilson2,3, Samantha MaWhinney2, Jane Reusch4,5,6, Leslie Knaub4,5, Sara Hull4,5, and Kristine M. Erlandson7,8
Abstract
Background
Muscle mitochondrial dysfunction associated with HIV and antiretroviral therapy (ART) may improve with exercise.
Methods
Muscle specimens obtained before and after 24 weeks of exercise in older PWH (n=18; ART >2 years) and uninfected controls (n=21) were analyzed for citrate synthase (CS) activity and complexes (C) I-V, manganese superoxide dismutase (MnSOD), peroxisome proliferator-activated receptor-γ coactivator-1 (PGC1α), and voltage-dependent anion channel 1 (VDAC1) content.
Results
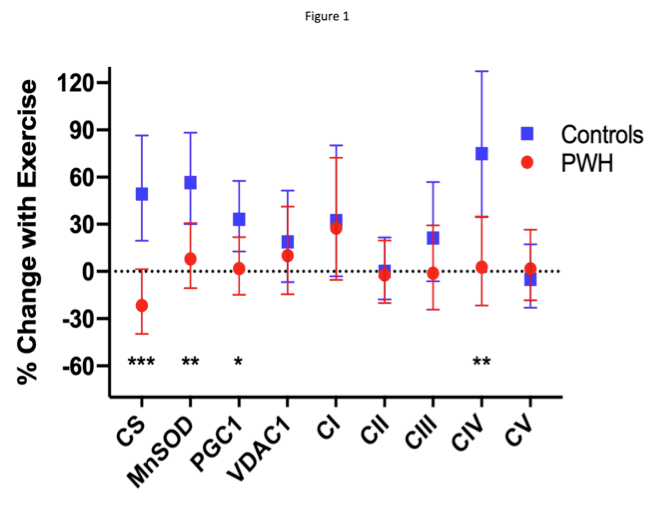
Only controls had increased CS, MnSOD, PGC1 and CIV (P≤0.01; P< 0.01 vs PWH) after training.
Conclusions
The blunted mitochondrial adaptations to training in PWH suggests the need for different types of exercise-induced stimulation.
Background
Aging, HIV infection, and antiretroviral therapy (ART) are associated with mitochondrial damage and dysfunction in skeletal muscle [1-4]. In muscle, lower total mitochondrial DNA [2], enzyme activity (e.g., citrate synthase), and mitochondrial respiration [5] have been associated with older age, lower aerobic capacity [2], reduced physical function [5], and a sedentary lifestyle during older age [6]. In contrast, exercise is a potent stimulus for the regulation of skeletal muscle mitochondrial bioenergetics, biogenesis, and remodeling [7] and a physically active lifestyle during older age may preserve or reverse mitochondrial function [6].
PWH experience earlier manifestations of muscle impairment, including greater than expected impairments in physical function [8, 9] than uninfected peers. We found that, when compared to uninfected controls, older PWH had worse baseline physical function [10] and greater improvements in cardiorespiratory capacity (VO2 peak; 400-m walk) after 24 weeks of exercise training. Given the results of our exercise training study in older PWH [10] and the deleterious effects of HIV infection, ART, aging, and a sedentary lifestyle on mitochondrial function, we hypothesized lower muscle mitochondrial content and enzyme activity in older PWH than uninfected controls, and HIV-specific changes in these markers in response to exercise training .
Discussion
We found HIV-specific differences in muscle mitochondrial markers at baseline and, most notably, blunted adaptations to exercise training in older men with decades of HIV and ART exposure. The intervention included moderate- to high-intensity cardiovascular endurance and resistance exercise to align with the physical activity guidelines for the general U.S. population and for PWH [11].
At baseline (pre-training), PWH had greater VDAC-1 content and lower electron transport complex III and IV content compared to controls. To the best of our knowledge, we are the first to report on HIV-specific differences in VDAC-1 content in skeletal muscle. VDAC-1 (mitochondrial porin), is a protein located on the outer mitochondrial membrane that selectively regulates the exchange of calcium ions, ATP, and other metabolites between the cytoplasm and the inner mitochondria, and also regulates apoptosis [13]. VDAC-1 may be elevated as the result of apoptotic stimuli in the cytoplasm that trigger increases in mitochondrial ionized calcium levels, further inducing the apoptotic cascade [13]. Whether this apoptotic mechanism is heightened in muscle exposed to HIV and ART, or is an adaptation resulting from greater mortality risk (VACS-1) is to be determined.
Reduced oxidative capacity and electron transport function in sedentary PWH at baseline was suggested by a trend for lower CS activity (33%; P=0.079) and significantly lower content of electron transporter complex III and IV subunits when compared to controls. Notably, the PWH in this study also had significantly slower walk times at baseline than controls [10], a functional outcome consistent with lower energy production in muscles of ambulation, deleterious effects of HIV infection and aging [5], and sedentarism [6]. Our findings align with Ortmeyer et al [14] who found that CS activity was 77% lower in middle- aged men with ART-treated HIV compared to controls, although they found no HIV-specific differences in Complexes I-V in muscle tissue.
The effects of exercise training on mitochondria diverged in PWH and controls. The impaired mitochondrial adaptations to exercise training in older PWH encompassed a wide scope of functions including substrate delivery and energy generation, oxidative capacity, antioxidant defense, and mitochondrial biogenesis and networking.
Our findings of the effects of exercise on the mitochondrial markers among controls were consistent with Irving et al [15] who found trends for increased skeletal muscle PGC1 and CS activity after 8 weeks of combined moderate-intensity aerobic and resistance exercise training in older adults. As shown in our adjusted models, the increased CS activity and content of MnSOD, PGC-1α, and CIV in controls (but not PWH) could not be attributed to better exercise adherence or differences in body composition, with the exception of adiposity moderating the changes in MnSOD. The increase in VO2 peak in controls [10] could be attributed to an increase in CS activity. In older men with long-standing HIV infection, most with previous exposure to thymidine analogs, the increase in VO2 peak without improvements in mitochondrial function suggests alternative adaptive pathways and long-term limitations on mitochondrial function. It is not known whether other exercise strategies, such as high-intensity interval training, could elicit improvements in mitochondrial function that would further increase aerobic capacity and exercise tolerance in older PWH. We previously found [10] that PWH who exercised at a higher versus lower intensity tended o have greater improvements in physical function. In the present study, we did not detect significant effects of exercise intensity on the mitochondrial markers and were underpowered to detect an interaction of HIV serostatus with exercise intensity.
Although this study has several strengths, including a comparison of baseline and training-induced changes in mitochondrial markers in middle-aged PWH and uninfected controls, limitations are acknowledged. The exclusively male sample limits generalization to women aging with HIV. The exercise intensity was increased after week 12 for approximately half of PWH and controls but muscle biopsies were not obtained at this juncture. Because the muscle specimens had been frozen, mitochondrial respiration could not be measured. Finally, we cannot ascertain the relative contributions of HIV infection, use of current ART formulations, or prior exposure to thymidine nucleotides to the mitochondrial responses.
We found that current exercise recommendations for adults were not sufficient to induce mitochondrial adaptations in older PWH. Future studies with measures of muscle mitochondrial respiration are needed to further elucidate the pre-training differences and exercise responses that are HIV-specific. We contend that a different exercise strategy, such as high-intensity interval training, may be necessary to stimulate mitochondrial adaptations or alternative mechanisms in skeletal muscle that would improve exercise tolerance of people aging with HIV.
Results
Pre-training muscle biopsies were obtained from 18 male PWH and 21 controls (Table 1; cross-sectional cohort). PWH had a similar age, significantly lower BMI and percent body fat, slower 400-m walk time, and were more likely to have a greater comorbid burden (higher Veterans Aging Cohort Study score) than controls. PWH were well controlled on ART, as indicated by an HIV-1 RNA < 20 copies/mL in all participants and a median CD4 count over 500 cells/uL. Median time since HIV diagnosis and ART use were a decade or more, and the majority of PWH had been exposed to thymidine analogues.
In the 15 PWH and 16 controls with paired pre- and post-training muscle biopsies (Table 1, training cohort), PWH had lower BMI and percent body fat (P=0.001) and slower 400-m walk time (P=0.026) than controls. The baseline characteristics of the training and cross-sectional cohorts were not significantly different.
Pre-Training Comparisons by HIV Serostatus
In the pre-training mitochondrial comparisons (Suppl Table 1), PWH had significantly greater content of VDAC-1 and lower CIII and CIV than controls; no other markers differed significantly by HIV serostatus. After adjusting for percent body fat, only CIII remained significantly lower in PWH.
Exercise Training Comparisons by HIV Serostatus (Paired Samples)
At baseline, the mitochondrial characteristics of the training and cross-sectional cohorts were comparable except that CIII content was not significantly different between PWH and controls in the training cohort (Suppl Table 2). After 24 weeks of exercise training, CS activity, MnSOD, PGC-1, and CIV increased significantly among controls, with decreases (CS) or minimal increases (MnSOD, PGC-1, CIV) among PWH, resulting in significant differences by HIV serostatus (Fig 1, Suppl Table 3). These comparisons remained significant when additionally adjusting for age. When adjusted for percent body fat and pre- training levels, MnSOD changes were no longer different by HIV serostatus. Changes in the other mitochondrial markers did not differ significantly by HIV serostatus. The changes in the mitochondrial markers were not significantly different by exercise intensity (moderate- intensity versus high-intensity; data not shown). Finally, we report the effect of training on the relationship between CS activity and VO2 peak. When pooling all participants, the changes in VO2 peak and CS activity were inversely related (-5.6% (-10.7, -0.3)%; CS activity decreased per 1 unit increase in VO2 (P=0.04). When adjusting for change in VO2 peak and HIV status, controls had significantly greater changes in CS activity than PWH (P=0.04). Assuming the overall mean VO2 peak change of 4.4 ml/kg/min, significant changes in CS activity were seen in controls (43% (14, 79%); P=0.004) but not in PWH (-7% (-32, 26%); P=0.63. No other training-induced changes in mitochondrial markers were significantly related to changes in VO2 peak.
|
|
| |
| |
|
|
|