 |
|
|
| |
Clonally expanded HIV-1 proviruses with 5'-leader
defects can give rise to nonsuppressible residual viremia
|
| |
| |
Download the PDF here
Download the PDF here
In conclusion, we demonstrate that proviruses with 5'-L defects, when in favorable genomic and immunological conditions, are a frequent cause of persistent NSV despite effective ART. Although incapable of causing viral rebound, these proviruses contribute to variable levels of viremia and complicate ART management (84), leading to additional testing, unnecessary ART changes and intensifications, and causing anxiety and frustration in patients and clinical care providers. Our work reveals additional complexity in residual viremia and should prompt the development of assays that would allow detection of 5'-L defects.
January 5, 2023
Jennifer A. White, … , Janet D. Siliciano, Francesco R. Simonetti
Jennifer A. White,1 Fengting Wu,1 Saif Yasin,2 Milica Moskovljevic,1 Joseph Varriale,1 Filippo Dragoni,1 Angelica Camilo-Contreras,1 Jiayi Duan,1 Mei Y. Zheng,2 Ndeh F. Tadzong,2 Heer B. Patel,2 Jeanelle Mae C. Quiambao,2 Kyle Rhodehouse,1 Hao Zhang,3 Jun Lai,1 Subul A. Beg,1 Michael Delannoy,4 Christin Kilcrease,1 Christopher J. Hoffmann,1 Sebastien Poulin,5 Frederic Chano,5 Cecile Tremblay,6,7 Jerald Cherian,1 Patricia Barditch-Crovo,1 Natasha Chida,1 Richard D. Moore,1 Michael F. Summers,2,8 Robert F. Siliciano,1,8 Janet D. Siliciano,1 and Francesco R. Simonetti1
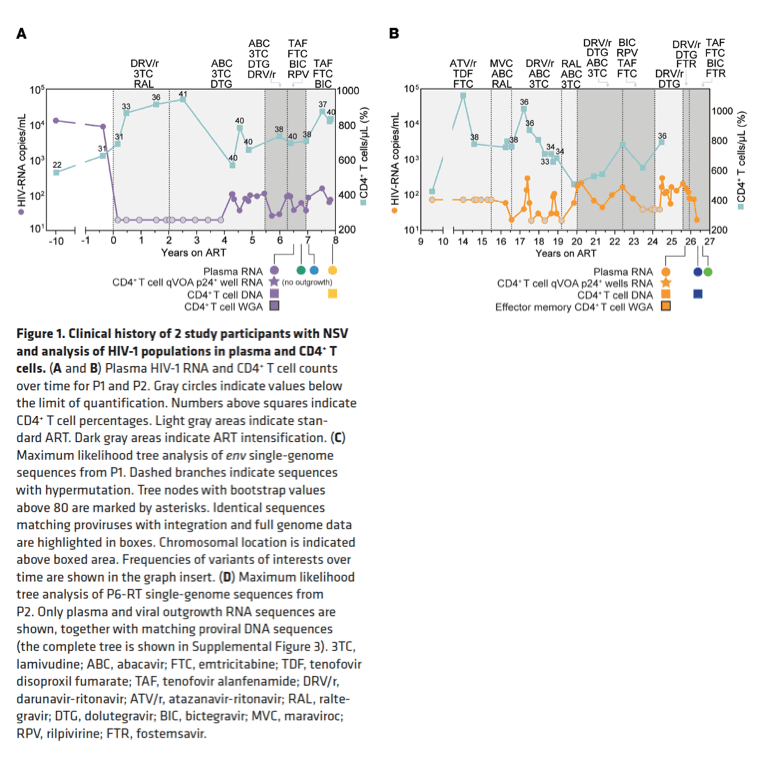
Emergence of NSV despite ART. Participants were referred for persistently detectable plasma viremia despite optimal adherence. Their characteristics and HIV-1 reservoir measurements are summarized in Supplemental Table 1, Figure 1, and Supplemental Figures 1 and 2 (supplemental material available online with this article; https://doi.org/10.1172/JCI165245DS1). The 4 participants had been living with HIV-1 for more than 15 years (range, 15-32 years) and had been on long-term ART (range, 7.8-27 years) to which they responded with peripheral CD4+ T cell recovery and undetectable viral loads. After years of viral suppression (<20 copies/mL), they experienced detectable viremia not explained by changes in adherence, drug bioavailability, or resistance. Treatment optimization, intensification, or deintensification had no effect (Figure 1, A and B, and Supplemental Figures 1 and 2). Participant 1 (P1) had persistently detectable viremia for 4.3 years, with a median of 80 HIV-1 RNA copies/mL of plasma (range, 37-156 copies/mL). P2 experienced intermittent periods of detectable viremia for more than 10 years, with a median of 75 copies/mL (range, <20-300 copies/mL). P3 had persistently detectable viremia for 4.6 years, with a median of 123 HIV-1 RNA copies/mL (range, 26-857 copies/mL) (Supplemental Figure 1). P4 represents an extreme example of NSV, maintaining a median of 2,979 copies/mL (range, 1,145-5,138 copies/mL) for almost 2 years (Supplemental Figure 2). Clinical histories are summarized in Supplemental Results.
Abstract
Background. Antiretroviral therapy (ART) halts HIV-1 replication, decreasing viremia to below the detection limit of clinical assays. However, some individuals experience persistent nonsuppressible viremia (NSV) originating from CD4+ T cell clones carrying infectious proviruses. Defective proviruses represent over 90% of all proviruses persisting during ART and can express viral genes, but whether they can cause NSV and complicate ART management is unknown.
Methods. We undertook an in-depth characterization of proviruses causing NSV in 4 study participants with optimal adherence and no drug resistance. We investigated the impact of the observed defects on 5'-leader RNA properties, virus infectivity, and gene expression. Integration-site specific assays were used to track these proviruses over time and among cell subsets.
Results. Clones carrying proviruses with 5'-leader defects can cause persistent NSV up to approximately 103 copies/mL. These proviruses had small, often identical deletions or point mutations involving the major splicing donor (MSD) site and showed partially reduced RNA dimerization and nucleocapsid binding. Nevertheless, they were inducible and produced noninfectious virions containing viral RNA, but lacking envelope.
Conclusion. These findings show that proviruses with 5'-leader defects in CD4+ T cell clones can give rise to NSV, affecting clinical care. Sequencing of the 5'-leader can help in understanding failure to completely suppress viremia.
Funding. Office of the NIH Director and National Institute of Dental and Craniofacial Research, NIH; Howard Hughes Medical Institute; Johns Hopkins University Center for AIDS Research; National Institute for Allergy and Infectious Diseases (NIAID), NIH, to the PAVE, BEAT-HIV, and DARE Martin Delaney collaboratories.
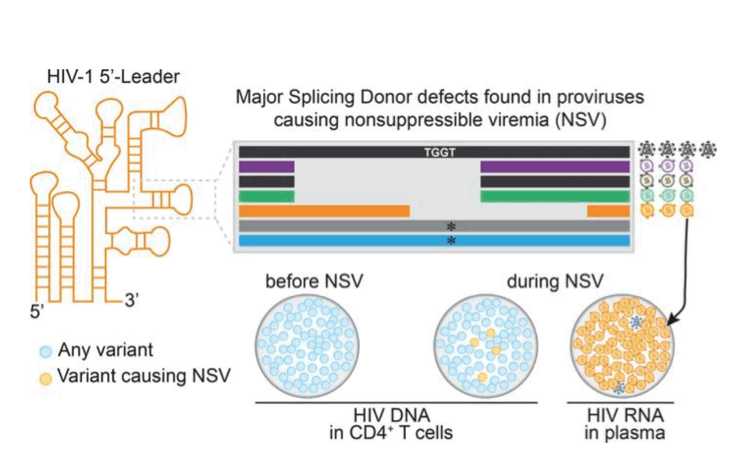
INTRODUCTION
Treatment with antiretroviral therapy (ART) rapidly reduces plasma HIV-1 to below the detection limit of clinical assays, prevents infection of new cells, and dramatically reduces HIV-1-associated morbidity and mortality (1). However, ART is not curative because the virus persists in a stable latent reservoir in resting memory CD4+ T cells, necessitating life-long adherence to ART (2-5).
A small fraction of reservoir cells become activated daily and produce trace levels of free virus (1-3 copies/mL) detectable with ultrasensitive assays (6, 7). These viruses are nonevolving archival sequences sensitive to the current regimen (8-10), indicating that residual viremia reflects virus release from stable reservoirs rather than ongoing replication cycles. Residual viremia cannot be suppressed by treatment intensification (11, 12) and is often dominated by identical sequences reflecting extensive proliferation of infected T cells (10, 13-15), which contributes to reservoir stability (16-20).
Some treated people living with HIV-1 (PLWH) experience sustained periods of detectable viremia, prompting reassessment of adherence, drug resistance, and drug concentrations. Regimen optimization or intensification often fails to suppress viremia (11, 12, 21). Recent studies have shown that expanded T cell clones carrying infectious proviruses can cause nonsuppressible viremia (NSV) (14, 15).
In treated PLWH, more than 90% of proviruses are defective (22, 23) with large deletions and/or APOBEC3G/F-induced G-to-A hypermutations (24). Some defective proviruses can express HIV-1 RNA and produce viral proteins (25-30). A recent study suggested that a fraction of viruses found in plasma before ART was defective due to small deletions and frameshifts (31). Some defective proviruses have small deletions in the 5'-leader (5'-L) upstream of gag (23, 32, 33). This region contains regulatory elements orchestrating genomic RNA dimerization and packaging, reverse transcription, proviral gene expression, and RNA splicing and translation (34-39). Proviruses with small 5'-L deletions make up 5% to 10% of proviral populations, are found in most PLWH, often in expanded clones (23, 32, 40-42), and may produce viral mRNA and p24 protein (26, 33). Given the potential of 5'-L defects to affect multiple steps in the viral cycle, these proviruses would not be expected to cause NSV or viral rebound. However, we demonstrate that these proviruses are a common cause of NSV that complicates clinical management.
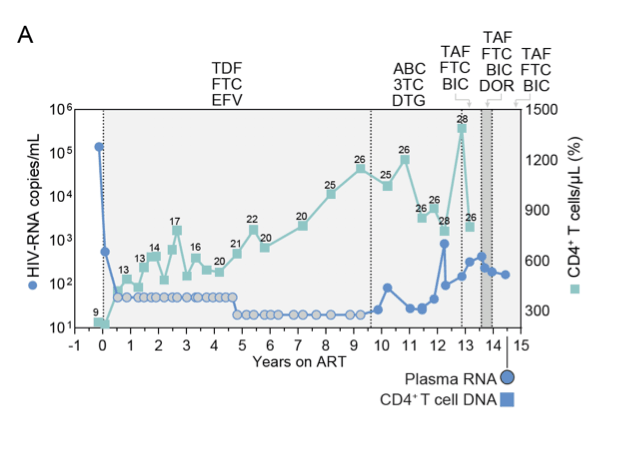
Supplementary Figure S1. Clinical history of Study Participant P3 with nonsuppressible viremia (NSV), and analysis of HIV-1 popu-lations in plasma and CD4+ T cells. (A) Plasma HIV-1 RNA and CD4+ T cell counts over time for P3; grey circles indicate values below the limit of quantification of the clinical assay; numbers above squares indicate CD4+ T cell percentage; light grey areas indicate standard ART, dark grey areas indicate ART intensification; ART regimens are indicated above the graph; time points and samples analyzed in this study are shown below the graph.
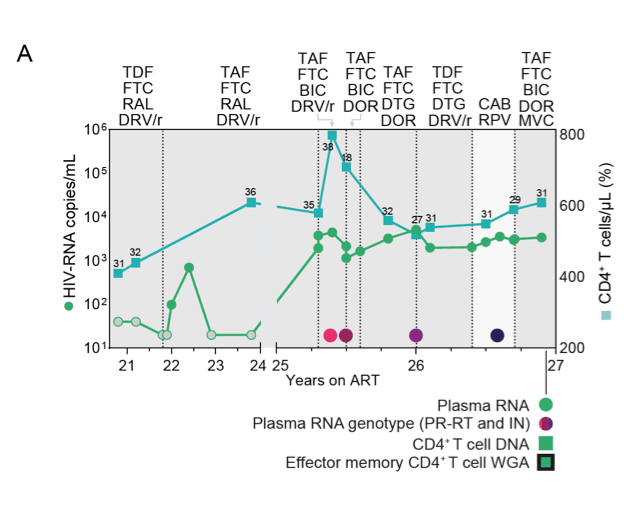
Supplementary Figure S2. Clinical history of study participant P4 with nonsuppressible viremia (NSV), and analysis of HIV-1 popula-tions in plasma and CD4+ T cells. (A) Plasma HIV-1 RNA and CD4+ T cell counts over time for P4; grey circles indicate values below the limit of quantification of the clinical assay; numbers above squares indicate CD4+ T cell percentage; light grey areas indicate standard ART, dark grey areas indicate ART intensification; ART regimens are indicated above the graph; time points and samples analyzed in this study are shown below the graph.
|
| |
|
|
|
|
|