| |
Effect of concurrent AZT use on the resistance pathway selected by abacavir-containing regimens
|
| |
| |
......Selection of both K65R and L74V/I was significantly reduced by co-administration of ZDV with ABC (one of 86 and two of 86 patients, respectively).... The L74V mutation causes decreased susceptibility to ddI...
HIV Medicine
November 2004
ER Lanier 1 , N Givens 2 , C Stone 3 , P Griffin 3 , D Gibb 4 , S Walker 4 , M Tisdale 3 , D Irlbeck 1 , M Underwood 1 , M St Clair 1 and M Ait-Khaled 2
1GlaxoSmithKline, Research Triangle Park, NC, USA, 2GlaxoSmithKline, Greenford, UK, 3GlaxoSmithKline, Stevenage, UK, and 4Medical Research Council Clinical Trials Unit, London, UK
INTRODUCTION
Abacavir (ABC) has been shown to select for four mutations in HIV-1 reverse transcriptase (RT) both in vitro and during monotherapy in vivo [1-3]. Individually, these mutations (K65R, L74V, Y115F and M184V) decrease susceptibility to ABC, but combinations of these mutations are required for >4.5-fold decreases in susceptibility to ABC, the clinically defined breakpoint for significant loss of ABC activity in vivo [4]. These four mutations cause varying levels of cross-resistance to other nucleoside or nucleotide reverse transcriptase inhibitors (NRTIs).
The K65R mutation is associated with broad cross-resistance to the NRTIs, reducing susceptibility to tenofovir (TDF), didanosine (ddI), zalcitabine (ddC), lamivudine (3TC), stavudine (d4T), emtricitabine (FTC) and amdoxovir (DAPD) in addition to ABC [5-11]. This mutation is associated with increased susceptibility to zidovudine (ZDV) [12]. The L74V mutation causes decreased susceptibility to ddI, ddC and ABC and increased susceptibility to TDF and ZDV [1,13,14]. The Y115F mutation is associated with reduced susceptibility to ABC and may be selected in vitro by TDF [1,15]. The M184V mutation reduces susceptibility to 3TC, FTC, ddC and ddI, but increases susceptibility to ZDV, d4T and TDF [14,16-19].
ABC does not select thymidine-analogue mutations (TAMs), associated with d4T and ZDV, either in vitro or during monotherapy in vivo. However, certain combinations of TAMs, most notably the M41L/L210W/T215Y complex, can cause broad cross-resistance to the NRTI class and have been shown to reduce ABC activity [4,20].
Previous studies have suggested that the pattern of resistance mutations selected by ABC-containing NRTI regimens differs according to the other NRTIs in the regimen. For example, ABC/3TC with or without nelfinavir (NFV) appears to select for the same four mutations seen with ABC monotherapy [21]. In contrast, ABC/ZDV is associated primarily with wild-type virus at rebound [2,21]. The triple therapy regimen ABC/3TC/ZDV selects initially for the M184V mutation, followed by TAMs in some cases [22,23]. The K65R, L74V and Y115F mutations are uncommon with this combination.
We performed a retrospective analysis of the resistance mutations observed in virus samples from patients in whom HIV-1 replication was inadequately suppressed on ABC-containing therapy.
ABSTRACT
Objectives: Abacavir (ABC) selects for four mutations (K65R, L74V, Y115F and M184V) in HIV-1 reverse transcriptase (RT), both in vitro and during monotherapy in vivo. The aim of this analysis was to compare the selection of these and other nucleoside reverse transcriptase inhibitor (NRTI)-associated mutations by ABC-containing therapies in the presence and absence of concurrent lamivudine (3TC) and/or zidovudine (ZDV) and to assess the effect of these mutations on phenotypic susceptibility to the NRTIs. Design: This study was a retrospective analysis of the patterns of NRTI-associated mutations selected following virological failure in six multicentre trials conducted during the development of ABC. Methods: Virological failure was defined as confirmed vRNA above 400 HIV-1 RNA copies/mL. RT genotype and phenotype were determined using standard methods. Results: K65R was selected infrequently by ABC-containing regimens in the absence of ZDV (13 of 127 patients), while L74V/I was selected more frequently (51 of 127 patients). Selection of both K65R and L74V/I was significantly reduced by co-administration of ZDV with ABC (one of 86 and two of 86 patients, respectively). Y115F was uncommon in the absence (seven of 127 patients) or presence (four of 86 patients) of ZDV. M184V was the most frequently selected mutation by ABC alone (24 of 70 patients) and by ABC plus 3TC (48 of 70 patients). Thymidine analogue mutations were associated with ZDV use. The K65R mutation conferred the broadest phenotypic cross-resistance of the mutations studied. Conclusions: The resistance pathway selected upon virological failure of ABC-containing regimens is significantly altered by concurrent ZDV use, but not by concurrent 3TC use. These data may have important implications for the efficacy of subsequent lines of NRTI therapies.
AUTHOR DISCUSSION
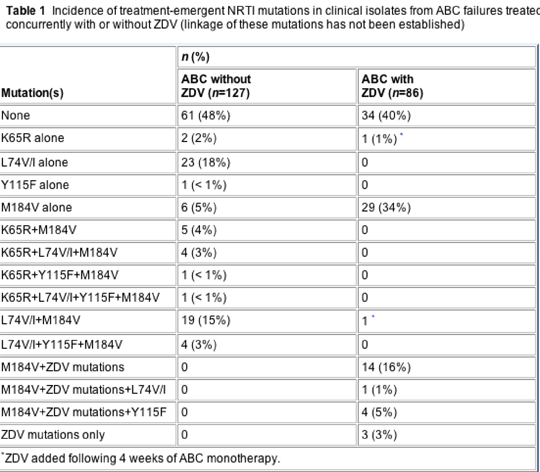
These results confirm and extend previous observations that concurrent use of thymidine analogues may reduce the incidence of K65R selection [30-33]. In this study, K65R was detected in only one of 86 samples from subjects on a failing ABC regimen that included ZDV, and that patient had received ABC monotherapy prior to the addition of ZDV (CNA2001). K65R by comparison was detected in 13 of 127 samples from patients failing ABC-containing regimens that did not include ZDV. The incidence of L74V was also significantly reduced by concurrent ZDV treatment (from 40% to 2%; Table 1). It is noteworthy that the therapy-experienced subjects in CNA2007 had a high prevalence of pre-existing TAMs, no selection for K65R in the absence of ZDV, but a very high rate of selection for L74V (53%). This suggests that the TAM/K65R pathways may be antagonistic, but that ZDV may be required to reduce L74V emergence. Selection for M184V on ABC+ZDV in the absence of 3TC was uncommon, being detected in one sample from a subject in CNA2001 where ABC monotherapy preceded addition of ZDV.
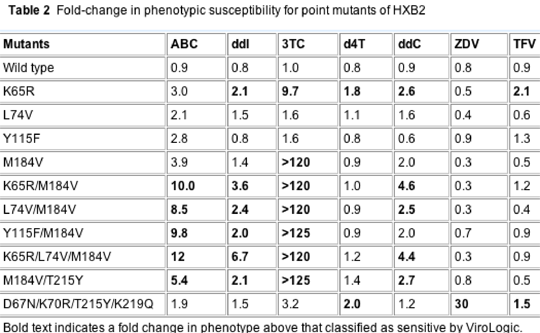
Given that M184V, L74V and K65R increase phenotypic susceptibility to ZDV in vitro, individually or in combination (Table 2), it is possible that ZDV is also more effective in vivo against virus with these mutations, reducing the likelihood that they will emerge in the dominant population. The mechanism behind the increased phenotypic susceptibility of HIV with these mutations to ZDV is not entirely clear, but may involve decreased ability of RT with these mutations to remove ZDV following incorporation [12,34,35]. The preponderance of wild-type RT in virological failures on ABC+ZDV±NFV (23 of 29 patients) from CNA2001 and PENTA 5 suggests a high genetic barrier to resistance for this combination, consistent with the TAM pathway selected in some patients during virological failure with ABC/3TC/ZDV [22,23,29].
Previous studies have suggested that K65R is negatively associated with thymidine analogue (ZDV and d4T) use [30-33]. However, recent studies have shown that d4T can select for K65R in vitro and that this mutation can cause decreased susceptibility to d4T, consistent with the phenotypic data presented here [9]. Additionally, K65R has emerged following virological failure on several d4T-containing regimens [36,37]. These data suggest that d4T may be less useful as a resistance modulator than ZDV, at least for K65R. Given the rapidly increasing prevalence of K65R (e.g. 0.8% of all samples in 1998 to 3.6% in 2003 for the Virco database [40]), several recent reports of high failure rates in regimens selecting this mutation and the resistance implications, understanding the factors involved in its selection may be clinically important [38-40].
It is important to note that most of these studies involved suboptimal regimens (mono or dual NRTIs) or highly experienced patients (CNA2007) and frequently long periods of detectable viraemia prior to the last genotype (reported here), so these results probably reflect worst case scenarios for mutation selection. Recent studies with ABC/3TC in conjunction with various third agents have found very low rates of selection for K65R or L74V shortly after virological failure [37,41].
The results of this study demonstrate that ZDV is useful as a resistance modulator, at least in combination with ABC, effectively reducing the incidence of selection for K65R and L74V. This may increase the likelihood that agents such as ddI, TDF and others affected by these two mutations will remain efficacious in second-line regimens. The relevance of these findings to combinations including NRTIs other than ABC, 3TC and ZDV remains to be elucidated.
RESULTS
The incidences of treatment-emergent NRTI mutations after virological failure on ABC-containing regimens with concurrent ZDV (n=86) and without concurrent ZDV (n=127) are shown in Table 1. Selection of K65R was infrequent (13 of 127 patients; 10%) following virological failure with ABC monotherapy (CNA2001 and CNA2002) or ABC/3TC±NFV (PENTA 5) in previously ART-naïve patients or ABC/EFV/APV (CNA2007) in ART-experienced patients, while L74V or I was selected more frequently (51 of 127 patients; 40%). In the monotherapy studies (treatment duration 12-24 weeks), M184V was the most common mutation detected (24 of 70 patients; 34%), followed by L74V (14 of 70 patients; 20%), K65R (10 of 70 patients; 14%) and Y115F (three of 70 patients; 4%). No mutations were detected for 32 of 70 patients (46%) in the monotherapy studies. ABC/3TC±NFV selected for M184V in eight of eight virological failures in PENTA 5 (treatment duration 48 weeks); in six of these eight failures, L74V was also detected, and in five of these six K65R and/or Y115F (one subject had K65R/L74V/Y115F and M184V) was detected [21]. Notably, K65R was not selected during 48 weeks of treatment for CNA2007, in which most patients (45 of 49; 92%) had baseline virus with TAMs; however, L74V or I emerged for 26 (53%) of these 49 patients and an additional six had L74V or I at baseline in the context of TAMs. Y115F was selected in two of 49 subjects (4%) and M184V in one of 49 subjects (2%) in CNA2007; 55% had M184V at baseline and one patient had Y115F at baseline [24].
|
|
| |
| |
 |
|
| |
| |
Selection of both the K65R and L74V or I mutations was significantly lower (P<0.001) when ZDV was co-administered with ABC. Only three of 86 samples from subjects failing a ZDV/ABC-containing regimen had K65R or L74V and two of these were from CNA2001, where ZDV was added after 4 weeks of ABC monotherapy. Y115F was uncommon in the absence (seven of 127 patients; 6%) or presence (four of 86 patients; 5%) of ZDV. The M184V mutation was observed for one patient on ABC+ZDV in the absence of 3TC in CNA2001, where ZDV was added after 4 weeks of ABC monotherapy. M184V was the most common mutation during ABC monotherapy (24 of 70 patients; 34%) or when 3TC was present (48 of 70 patients; 69%).
Overall, the incidence of M184V in the absence of ZDV was 31% versus 56% in the presence of ZDV treatment. This difference reflects the relative usage of 3TC in each group and selection for M184V by ABC alone. In the group that did not receive ZDV, only 6% (eight of 127 patients) also received 3TC. In the group that did receive ZDV, 66% (57 of 86 patients) also received 3TC. TAMs were associated with ZDV use, but appeared slowly when detected, as previously described for ABC/3TC/ZDV [22,23,29]. No TAMs were detected during 12 weeks of treatment (4 weeks of ABC monotherapy followed by 8 weeks of ABC/ZDV) in the ZDV-containing group of CNA2001 (16 of 18 wild-type) and four of 11 subjects developed TAMs during 48 weeks of treatment in the ABC/ZDV±NFV group from PENTA 5, with only wild-type virus detected in the other seven patients.
The effects of some of the treatment-emergent mutational patterns on phenotypic susceptibility were evaluated for a series of point mutants in an HXB2 background (Table 2). Changes in phenotypic susceptibility are shown as a ratio of the Virologic WT control (PNL4-3) to the test virus. Fold-changes that result in phenotypic susceptibility above that and are classified as susceptible by Virologic are shown in bold in Table 2, and may indicate reduced activity of the drug for viruses with the mutation in question.
|
|
| |
| |
 |
|
| |
| |
Of the four mutations associated with ABC monotherapy (K65R, L74V, Y115F and M184V), only the K65R mutation conferred broad cross-resistance when present in isolation. None of these mutations alone conferred 4.5-fold decreased susceptibility to ABC, the clinically defined breakpoint. The M184V mutation together with any other ABC-associated mutation was associated with reduced susceptibility to ABC, ddI, 3TC and ddC. As expected from previous studies, M184V and L74V increased phenotypic susceptibility to TDF [14]. The ABC-associated mutations (except Y115F) increased susceptibility to ZDV. The M184V mutation appeared to restore full susceptibility to d4T in the presence of K65R and partially restore susceptibility to TDF.
Materials and methods
We analysed virus samples from subjects who experienced virological failure in six studies conducted during the development of ABC. Virological failure genotypes from ABC monotherapy (CNAA2001, n=27; CNAB2002, n= 43) and combination therapies not containing ZDV (PENTA 5, ABC+3TC±NFV, n=8; CNAA2007, ABC+EFV+APV, n=49) were compared with those from ABC+ZDV-containing regimens (CNAA2001, ABC monotherapy for 4 weeks, followed by 8 weeks of ABC+ZDV, n=18; PENTA 5, ZDV+ABC±NFV, n=11; CNA3005, ABC+3TC+ZDV, n=40; and CNAA/B3003, ABC+3TC +ZDV, n=17). Genotypes reported are the last available while patients were on originally randomized therapy (CNA2001, week 12; CNA2002, median of week 20; PENTA 5, week 48; CNAA2007, median of week 48; CNA3003, week 48; CNA3005, median of week 72).
All of these studies were conducted in antiretroviral therapy (ART)-naïve subjects with the exception of CNAA2007. Detailed descriptions of these trials have been previously published [2,3,21-26]. Mutations known to be associated with decreased phenotypic sensitivity to ddC, d4T, ddI, 3TC, ZDV, TDF and/or ABC were analysed at HIV-1 RT codons 41, 62, 65, 67, 69, 70, 74, 75, 77, 115, 116, 151, 184, 210, 215 and 219 [27]. The incidence of the ABC-associated mutations K65R, L74V, Y115F and M184V was compared with the incidence of these mutations in ABC regimens without ZDV following virological failure using a chi 2 test with a significance level of P<0.01.
Site-directed mutagenesis was carried out using the QuikChange Mutagenesis Kit (Stratagene, La Jolla, CA) according to the manufacturer's protocol. Assays of phenotypic susceptibility to the marketed NRTIs were conducted by Virologic (South SanFrancisco, CA, USA) using PhenoSenseTM as described [28].
REFERENCES
1 Tisdale M, Alnadaf T, Cousens D et al. Combination of mutations in human immunodeficiency virus type 1 reverse transcriptase required for resistance to the carbocyclic nucleoside 1592U89. Antimicrob Agents Chemother 1997; 41: 1094-1098.
2 Harrigan P, Stone C, Griffin P et al. Resistance profile of the human immunodeficiency virus type 1 reverse transcriptase inhibitor abacavir (1592U89) after monotherapy and combination therapy. J Infect Dis 2000; 181: 912-920.
3 Miller V, Ait-Khaled M, Stone C et al. HIV-1 reverse transcriptase (RT) genotype and susceptibility to RT inhibitors during abacavir monotherapy and combination therapy. AIDS 2000; 14: 163-171.
4 Lanier ER, Ait-Khaled M, Scott J et al. Antiviral efficacy of abacavir in antiretroviral therapy-experienced adults harbouring HIV-1 with specific patterns of resistance to nucleoside reverse transcriptase inhibitors. Antiviral Ther 2004; 9: 37-45.
5 Wainberg MA, Miller MD, Quan Y et al. In vitro selection and characterization of HIV-1 with reduced susceptibility to PMPA. Antiviral Ther 1999; 4: 87-94.
6 Winters MA, Schafer RW, Jellinger RA et al. Human immunodeficiency virus type 1 reverse transcriptase genotype and drug susceptibility changes in infected individuals receiving dideoxyinosine monotherapy for 1-2 years. Antimicrob Agents Chemother 1997; 41: 757-762.
7 Zhang D, Caliendo AM, Eron JJ et al. Resistance to 2',3'-dideoxycytidine conferred by a mutation in codon 65 of the human immunodeficiency virus type 1 reverse transcriptase. Antimicrob Agents Chemother 1994; 38: 282-287.
8 Gu Z, Gao Q, Fang H et al. Identification of a mutation at codon 65 in the IKKK motif of reverse transcriptase that encodes human immunodeficiency virus resistance to 2',3'-dideoxycytidine and 2',3'-dideoxy-3'-thiacytidine. Antimicrob Agents Chemother 1994; 38: 275-281.
9 Garcia-Lerma J, MacInnes H, Bennett D et al. A novel genetic pathway of human immunodeficiency virus type 1 resistance to stavudine mediated by the K65R mutation. J Virol 2003; 77: 5685-5693.
10 Mulamba GB, Rimsky L, Myrick F, Borroto-Esoda K, Feng JY. Pre-steady state kinetic analysis of the incorporation of FTC 5'-monophosphate and 3TC 5'-monophosphate by mutant HIV-1 RTs K65R, K65R/Q151M and Q151M. 16th International Conference on Antiviral Research. Savannah, GA, 27 April-1 May 2003 [Abstract 39].
11 Bazmi HZ, Hammond JL, Cavalcanti SC, Chu CK, Schinazi RF, Mellors JW. In vitro selection of mutations in the human immunodeficiency virus type 1 reverse transcriptase that decrease susceptibility to (-)-beta-D-dioxolane-guanosine and suppress resistance to 3'-azido-3'-deoxythymidine. Antimicrob Agents Chemother 2000; 44: 1783-1788.
12 Parikh U, Koontz D, Hammond J et al. K65R: a multi-nucleoside resistance mutation of low but increasing frequency. 12th International HIV Drug Resistance Workshop. Los Cabos, Mexico, June 2003 [Abstract 136].
13 St. Clair M, Martin JL, Tudor-Williams G et al. Resistance to ddI and sensitivity to AZT induced by a mutation in HIV-1 reverse transcriptase. Science 1991; 253: 1557-1559.
14 Harrigan P, Miller M, McKenna P, Brumme Z, Larder B. Phenotypic susceptibilities to tenofovir in a large panel of clinically derived human immunodeficiency virus type 1 isolates. Antimicrob Agents Chemother 2002; 46: 1067-1072.
15 Stone C, Ait-Khaled M, Craig C, Tisdale M. HIV-1 reverse transcriptase (RT) mutations identified by in vitro selection with tenofovir (TDF) +/- abacavir (ABC) and TDF +/- lamivudine (3TC). XIth International HIV Drug Resistance Workshop. Seville, Spain, July 2002 [Abstract 44].
16 Tisdale M, Kemp S, Parry N, Larder B. Rapid in vitro selection of human immunodeficiency virus type 1 resistant to 3'-thiacytidine inhibitors due to a mutation in the YMDD region of reverse transcriptase. Proc Natl Acad Sci USA 1993; 90: 5653-5656.
17 Wainberg MA, Saloman H, Gu Z et al. Development of HIV-1 resistance to (-)2'-deoxy-3'-thiacytidine in patients with AIDS or AIDS-related complex. AIDS 1995; 9: 351-357.
18 Gao Q, Gu Z, Parniak M et al. The same mutation that encodes low-level human immunodeficiency type I resistance to 2',3'-dideoxyinosine and 2',3'-dideoxycytidine confers high-level resistance to the (-) enantiomer of 2',3'-dideoxy-3'-thiacytidine. Antimicrob Agents Chemother 1993; 37: 1390-1392.
19 Larder BA, Kemp SD, Harrigan PR. Potential mechanism for sustained antiretroviral efficacy of AZT-3TC combination therapy. Science 1995; 269: 696-699.
20 Miller V, Larder BA. Mutational patterns in the HIV genome and cross-resistance following nucleoside and nucleotide analogue drug exposure. Antiviral Ther 2001; 6: 25-44.
21 Gibb D, Walker A, Kaye S et al. Evolution of antiretroviral phenotypic and genotypic drug resistance in antiretroviral-naive HIV-1-infected children treated with abacavir/lamivudine, zidovudine/lamivudine or abacavir/zidovudine, with or without nelfinavir (the PENTA 5 trial). Antiviral Ther 2002; 7: 293-303.
22 Ait-Khaled M, Rakik A, Griffin P, Gould J, Cutrel A, Fischl M. The CNA3003 international Study Group. Viral rebound during 48 wks of ABC/3TC/ZDV combination therapy in antiretroviral naïve adults is infrequent with emergent virus showing a limited number of resistance mutations. Antiviral Ther 1999; 4 (Suppl. 1): 52.
23 Melby T, Tortell S, Thorborn D et al. Time to appearance of NRTI-associated mutations and response to subsequent therapy for patients on failing ABC/COM. 8th Conference on Retroviruses and Opportunistic Infections. Chicago, IL, February 2001 [Abstract 448].
24 Ait-Khaled M, Rakik A, Griffin P et al. HIV-1 reverse transcriptase and protease resistance mutations selected during 16-72 weeks of therapy in isolates from antiretroviral therapy-experienced patients receiving abacavir/efavirenz/amprenavir in the CNA2007 study. Antiviral Ther 2003; 8: 111-120.
25 Ait-Khaled M, Rakik A, Griffin P et al. Mutations in HIV-1 reverse transcriptase during therapy with abacavir, lamivudine and zidovudine in HIV-1-infected adults with no prior antiretroviral therapy. Antiviral Ther 2002; 7: 43-51.
26 Staszewski S, Keiser P, Montaner J et al. Abacavir-lamivudine-zidovudine vs indinavir-lamivudine-zidovudine in antiretroviral-naïve HIV-infected adults. J Am Med Assoc 2001; 285: 1155-1163.
27 Hirsch MS, Brun-Vezinet F, Clotet B et al. Antiretroviral drug resistance testing in adults infected with human immunodeficiency virus type 1: 2003 recommendations of an International AIDS Society-USA Panel. Clin Infect Dis 2003; 37: 113-128.
28 Petropoulos C, Parkin N, Limoli K et al. A novel phenotypic drug susceptibility assay for human immunodeficiency virus type 1. Antimicrob Agents Chemother 2000; 44: 920-928.
29 Matheron SDD, Boue F, Livrozet JM et al. Triple nucleoside combination zidovudine/lamivudine/abacavir versus zidovudine/lamivudine/nelfinavir as first-line therapy in HIV-1-infected adults: a randomized trial. Antiviral Ther 2003; 8: 163-171.
30 Winston A, Mandalia S, Pillay D, Gazzard B, Pozniak A. The prevalence and determinants of the K65R mutation in HIV-1 reverse transcriptase in tenofovir-naive patients. AIDS 2002; 16: 2087-2089.
31 Winston A, Pozniak A, Gazzard B, Nelson M. Which nucleoside and nucleotide backbone combinations select for the K65R mutation in HIV-1 reverse transcriptase? 12th International HIV Drug Resistance Workshop. Los Cabos, Mexico, June 2003 [Abstract 137].
32 MacArthur RD, Crane LR, Alvarez D, Fairfax M, Richmond D, Curtis G. Factors associated with selection of the K65R mutation: a retrospective chart review. 2nd International AIDS Society Conference on Pathogenesis and Treatment. Paris, France, July 2003 [Abstract 835].
33 Ruane P, Luber A, Akil B et al. Factors influencing selection of K65R mutation among patients receiving tenofovir (TDF) containing regimens. 2nd International AIDS Society Conference on Pathogenesis and Treatment. Paris, France, July 2003 [Abstract 582].
34 Ray A, Basavapathruni A, Anderson K. Mechanistic studies to understand the progressive development of resistance in human immunodeficiency virus type 1 reverse transcriptase to abacavir. J Biol Chem 2002; 277: 40479-40490.
35 Meyer PR, Matsuura SE, Zonarich D et al. Relationship between 3'-azido-3'-deoxythymidine resistance and primer unblocking activity in foscarnet-resistant mutants of human immunodeficiency virus type 1 reverse transcriptase. J Virol 2003; 77: 6127-6137.
36 Roge B, Katzenstein T, Obel N et al. K65R with and without S68: a new resistance profile in vivo detected in most patients failing abacavir, didanosine and stavudine. Antiviral Ther 2003; 8: 173-182.
37 Lanier ER, Irlbeck D, Liao Q et al. Emergence of resistance-associated mutations over 96 weeks of therapy in subjects initiating ABC/3TC + d4T, EFV or APV/r. 43rd Interscience Conference on Antimicrobial Agents and Chemotherapy. Chicago, IL, September 2003 [Abstract H-910].
38 Farthing C, Khanlou H, Yeh V. Early virologic failure in a pilot study evaluating the efficacy of once daily abacavir (ABC), lamivudine (3TC) and tenofovir DF (TDF) in treatment naïve HIV-infected patients. 2nd International AIDS Society Conference on Pathogenesis and Treatment. Paris, France, July 2003 [Abstract 43].
39 Gallant JE, Rodriguez AE, Weinberg W et al. Early non-response to tenofovir DF (TDF)+abacavir (ABC) and lamivudine (3TC) in a randomized trial compared to efavirenz (EFV)+ABC and 3TC. ESS30009 unplanned interim analysis. 43rd Interscience Conference on Antimicrobial Agents and Chemotherapy. Chicago, IL, 14-17 September 2003 [Abstract H-1722a].
40 Bacheler L, Vermeiren H, McKenna P, Van Houtte M, LeCocq P. Trends in genotypic and phenotypic resistance among clinical samples, submitted for routine HIV resistance analysis. 43rd Interscience Conference on Antimicrobial Agents and Chemotherapy. Chicago, IL, 14-17 September 2003 [Abstract H-917].
41 MacManus S, Yates PJ, White S, Richards N, Snowden W. GW433908 in ART-naïve subjects: absence of resistance at 48 weeks with boosted regimen and APV-like resistance profile with unboosted regimen. 10th Conference on Retroviruses and Opportunistic Infections. Boston, MA, February 2003 [Abstract 598].
|
|
| |
| |
|
|
|