| |
Seminal Reservoirs during an HIV-1 Eradication Trial
|
| |
| |
Short Communication
AIDS RESEARCH AND HUMAN RETROVIRUSES
Volume 21, Number 9, 2005, pp. 768-775
GIUSEPPE NUNNARI, DANIELA LETO, JULIE SULLIVAN, YAN XU, KETTI E. MEHLMAN, JOSEPH KULKOSKY, and ROGER J. POMERANTZ
Institute for Human Virology and Biodefense, Thomas Jefferson University, Philadelphia, Pennsylvania 19107.
ABSTRACT
Despite dramatic reduction of the levels of human immunodeficiency virus type I (HIV-1) virions in blood and seminal plasma of infected patients, highly active antiretroviral therapy (HAART) does not eradicate HIV-1. Three patients, with less than 50 copies/ml of plasma viral RNA, were enrolled in this eradication protocol. Didanosine (DDI) and hydroxyurea (HU) were added to their baseline HAART and after a month of therapy, low dose OKT3, followed by a 2-week course of interleukin 2 (IL-2), was administrated. All antiretroviral therapy was then interrupted and the three patients developed viral rebound in the peripheral
blood.
The V3 loop region of the HIV-1 gp120 from cell-free viral RNA and proviral DNA in blood and seminal compartments was sequenced in one patient. The two major viral isolates in semen cells were macrophage-tropic (R5) and dual-tropic (R5X4), and these isolates were also present in the PBMCs.
Six months after the viral rebound, we demonstrated a shift toward dual tropism in semen cell-associated HIV-1 proviral DNA, with the first appearance of a T-lymphotropic (X4) provirus solely in this compartment. The virus responsible for the blood plasma viral rebound was never found in the semen microenvironment. This study suggests viral compartmentalization of the semen microenvironment after an intensification and stimulatory HIV-1 eradication protocol, with evidence of viral evolution.
BACKGROUND
THE INTRODUCTION OF HIGHLY ACTIVE ANTIRETROVIRAL THERAPY
(HAART) dramatically changed the natural history of HIV-1 infection.1,2 The drastic reduction to clinical undetectability (HIV-1 RNA levels 50 copies/ml) of blood plasma HIV-1 RNA viral load can now be obtained in many patients,1,2
with profound effects on mortality and morbidity.2 A parallel reduction of the HIV-1 viral load in genital fluids has also been demonstrated.3 Nonetheless, HAART does not lead to true viral eradication. Retroviral latency and continuous low levels of viral replication can still be detected not only as cell-free virus but also in different cell types of distinct compartments.4-9 Resting CD4 T-lymphocytes are considered the main known viral reservoir in HIV-1-infected individuals on virally suppressive HAART, and replication-competent virus can be recovered, in
vitro, from these provirus-positive cells.4,6 Recently, peripheral blood monocytes have also been shown to produce replicationcompetent virus.9
Viral latency and persistent or cryptic viral replication in specific microenvironments shielded by blood:tissue barriers, such as the central nervous system (CNS), retina, and testes, may take place and act as possible HIV-1 reservoirs on virally suppressive HAART.8 The presence of replication competent virus has been demonstrated in seminal cells, revealing not only the
active part played in the transmission of HIV-1, but also the potential involvement of this compartment in residual HIV-1 disease.7 This might represent an in vivo reservoir or sanctuary site, which may reinfect the peripheral bloodstream and lymphoid tissue, especially when HAART is discontinued, interfering with attempts at viral remission or eradication.
The use of HAART allowed the possibility of identifying HIV-1 reservoirs, which are now considered the main targets in approaching HIV-1 eradication. Based on our understandings of molecular lentiviral pathogenesis, different strategies can be developed to activate persistently infected cells, which might lead to virus-induced cell death and purging of the viral reservoirs.
In the present study, three patients were enrolled in an intensification and stimulatory translational eradication trial, based on the administration of didanosine (DDI) and hydroxyurea (HU), plus OKT3 and interleukin 2 (IL-2),10 followed by complete interruption of all antiretroviral therapy. HU was used, especially combined with DDI, with which it has synergistic effects in inhibiting HIV-1 replication.11
It was hypothesized that combining novel "intensification" therapy, with DDI/HU, and stimulation of latently infected cells with low-dose OKT3 plus IL-2, might lead to remission or eradication of HIV-1 in vivo in selected patients. The aim of the
present substudy was to evaluate the effects of this intensification and stimulatory approach targeting HIV-1 residual disease in a critical reservoir, the seminal microenvironment, to understand whether it could be affected by this therapeutic regimen, either directly through cell stimulation or indirectly leading to
possible reseeding of the seminal compartment.
Three HIV-1-infected men on virally suppressive HAART, with at least two prestudy plasma viral RNA determinations of less than 50 copies/ml, determined by clinical ultrasensitive reverse transcriptase polymerase chain reaction (RT-PCR) (Roche, Inc.), were recruited for this protocol, entitled "Residual
HIV-1 Disease Eradication (RHIDE)."10 All patients had stable baseline CD4 T-lymphocyte counts of greater than 500 cells/mm3 and no history of opportunistic infections or other significant medical conditions.
In brief, these patients were treated with novel "intensification" therapy using DDI and HU and "stimulation" therapy utilizing OKT3 (Orthoclone OKT3-muroMonab-CD3; Ortho- Biotech, Inc.) and IL-2 (Proleukin, Chiron, Inc.). When replication-competent virus from CD8-depleted peripheral blood mononuclear cells (PBMCs) was not obtainable in these patients from any site after the course of OKT3 and IL-2, then with agreement from the patient, stopping all anti-HIV-1 medications
was attempted with close monitoring to evaluate for potential viral "rebound." Each patient signed an Institutional Review Board (IRB)-approved consent form for this study. This study was also reviewed and approved by the Food and Drug Administration (FDA).
CD8 T-lymphocyte-depleted PBMCs and seminal cells from the patients were separately cocultured with phytohemagglutinin (PHA)-stimulated CD8 T-lymphocyte-depleted PBMCs from HIV-1-seronegative donors and monitored
for ex vivo HIV-1 growth for 6-10 weeks, as described previously.7 To determine levels of HIV-1 RNA in the blood plasma and seminal fluid, these samples were concentrated via ultracentrifugation at 35,000 rpm for 1 h, and RNA was isolated and analyzed by supersensitive RT-PCR, as described previously.7
Comparison of the test samples with this serially diluted standard curve of the amplified in vitro transcribed standard was utilized to quantitate viral unspliced RNA to 5 copies, within the linear amplification range of this assay. Of note, although outside the linear amplification range and therefore not amenable to precise quantitation, viral transcripts below 5 copies (i.e., 4 to 1 copy/ml) could nonetheless be detected using this assay system.7
The sequences of the V3 loop of the gp120 region of the viral envelope (env) gene of HIV-1 were determined by a nested PCR assay for proviral DNA and virion-encapsidated RNA. The V3 region was initially amplified by using an external primer pair KK30 and KK40, and internal primers KK10N and KK20N, which bear 5 EcoRI and 5 PstI sites, respectively, for unidirectional cloning into pUC19 digested with EcoRI and PstI. Sequencing of the positive clones was performed with an automated sequencer (Prism model 377 with XL Upgrade). The sequences have been submitted to GenBank, and the accession numbers are AY786499 to AY7864505. Perfectly matched DNA-based sequences, within each patient grouping, were not reiterated in the submission.
Sequence chromatogram data were analyzed using Seqman (DNASTAR, Madison, WI). Multiple alignments were performed with ClustalX version 1.83, utilizing the Gonnet series weight matrix and the IUB DNA weight matrix alignment parameters. Intrapatient viral envelope sequences were compared
to each other and to the control sequences (Bal1 and NL4-3), using Lasergene sequence analysis software obtained from DNASTAR Inc. to exclude any possibility of contamination. Bootstrap analysis consisting of 1000 replicates was performed with the ClustalX program. Bootstrap values of 70% or higher were considered significant.
No patient developed plasma viremia above 50 copies/ml during these therapies. In patient 1, plasma viral RNA became undetectable with stimulation and intensification therapy. As well, seminal fluid viral RNA remained undetectable (less than 1 copy/ml) (Table 1).
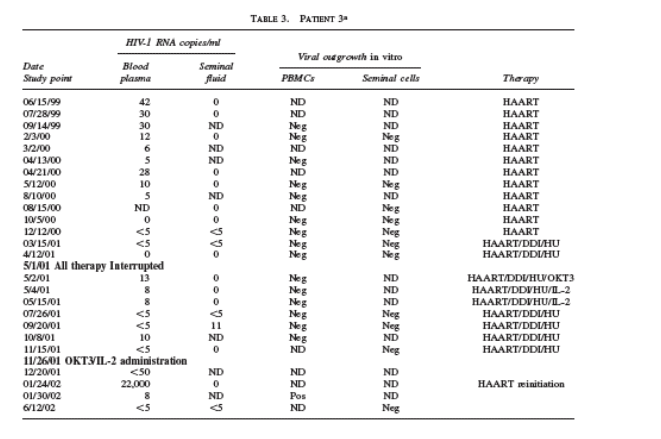
In patient 2, very low levels of plasma HIV-1 RNA were detectable at certain time points posttherapy with OKT3 plus IL-2 (Table 2). Seminal fluid HIV-1 RNA, which was undetectable before the initiation of the intensification and stimulatory
therapy, remained so at most time points and was detectable, but at 5 copies/ml (i.e., detectable but below quantitation) on three occasions (Table 2).
In patient 3, plasma and seminal viral RNA was able to be detected at several time points during this study, albeit at extremely low levels (Table 3). Importantly, seminal fluid HIV-1 RNA was undetectable (less than 1 copy/ml), when plasma
viral rebound occurred after antiretroviral therapy interruption. During the intensification and stimulatory therapy regimen, no ex vivo viral outgrowth was detected at any time points in patients 2 and 3, as measured by HIV-1 p24 antigen levels in culture supernatants of both CD8-depleted PBMCs and seminal
cell cocultures. In patient 1, no viral outgrowth was detected from seminal cells cocultured, even on the sporadic occasion when low levels of viral outgrowth could be detected from CD8-depleted PBMC cocultures.
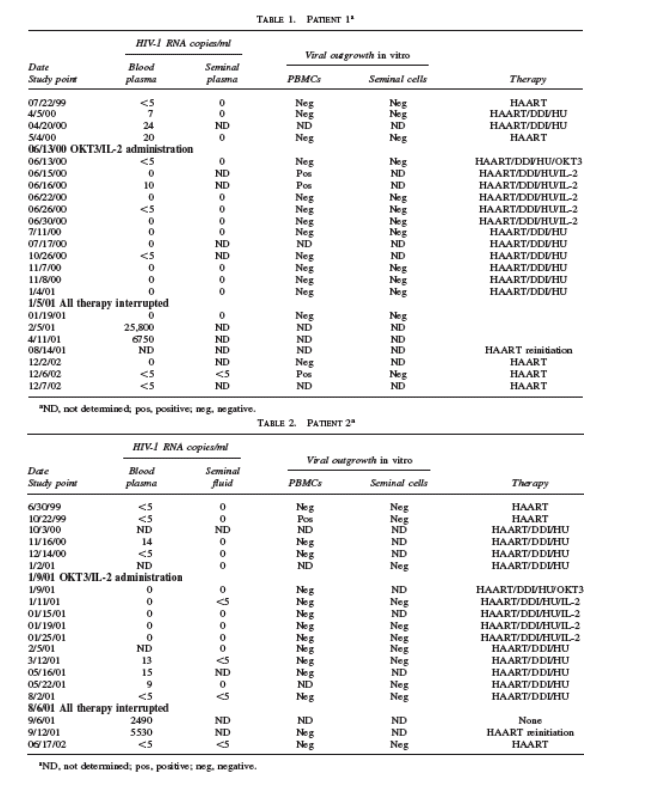
The sequences of the V3 loop region of HIV-1 gp120 of the viral envelope (env) from semen and blood compartments of patient 3 were analyzed and compared to determine the relationship between these two microenvironments before, during, and after the eradication approach. We were unable to sequence the V3 loop region of HIV-1 gp120 in the seminal compartments of patients 1 and 2 due to lack of sample availability, a sufficient number of cells, or HIV-1 DNA-positive samples. Moreover, when we further tried to detect HIV-1 proviral DNA gag copies in the semen cells of patients 1 and 2, determined by an ultrasensitive PCR technique able to detect to 5 copies of HIV-1 DNA/ml, we could not detect any positive sample (data not shown).
Two major viral isolates where detected in seminal cell-associated proviral DNA before intensification treatment, after OKT3 and IL-2 stimulation, and 6 months after the complete withdrawal of therapy (Table 4). Importantly, the proviruses
contained in the seminal reservoir did not appear to be the same quasispecies responsible for the blood plasma viral rebound, which occurred after interruption of therapy (Fig. 1). Six months after antiretroviral therapy was stopped, we detected a third viral DNA isolate in the seminal cells, a shift toward dual tropism, and the emergence of a T cell line-tropic (X4) virus. As shown in Fig. 1, semen isolates at poststudy evaluation dates were found on separate branches of the phylogenetic tree, suggesting seminal compartmentalization. Of importance,
the major virus detected during viral rebound, in peripheral blood, was never detected in the seminal compartment. The design of this clinical study was initially based on an "oncological paradigm" for treatment of HIV-1 infection.8 This includes effective HAART as "induction" therapy and then further approaches against HIV-1 latency, cryptic replication, and sanctuary sites for the removal of residual disease. Nonetheless, even when utilizing intensification therapy with DDI and HU added to baseline suppressive HAART, patients in which all antiretroviral therapy was stopped at the end of the protocol
developed plasma viral rebound.10
We further investigated whether this eradication approach could have had an impact on the seminal reservoir, either stimulating it as a potential source of viral rebound or reseeding it, especially when the patients underwent therapy interruption.
Relatively little is known regarding rebound viral strains in patients on virally suppressive HAART, when antiretroviral agents are stopped. A previous small study had suggested that rebound virus after stopping HAART may be present in PBMCs and lymphoid tissue.12 Another study had suggested that rebound virus may come from a nonblood source of yet undetermined reservoir sites,13 while a third study demonstrated that only actively replicating virus in PBMCs (i.e., cell-associated viral RNA) is the source of plasma rebound viral species.14
In this study, we analyzed seminal fluid HIV-1 RNA viral load and viral outgrowth of seminal cell cocultures over the course of the eradication trial. We found that all three parameters remained stable in all patients studied. In patient 3, when
blood plasma viral rebound occurred, seminal plasma HIV-1 viral levels remained undetectable, showing possible viral compartmentalization in the male genital tract.
Furthermore, in patient 3, seminal cell-associated proviruses were compared with viral isolates emerging in peripheral blood cells, cultured ex vivo, during stimulation therapy and, more importantly, with those responsible for viral plasma rebound.15 Based on viral sequences and tropism, we demonstrated that
the two major viral isolates in seminal cells were macrophagetropic (R5) and dual-tropic (X4R5), and these strains were also present in the PBMCs. Six months after the plasma viral rebound, we found a shift toward dual tropism in semen cell-associated HIV-1 proviral DNA, with the first appearance of a Tlymphotropic (X4) provirus solely in this compartment (Table 4). The virus responsible for the blood plasma viral rebound was not found in the semen microenvironment, either before or at completion of the protocol. Thus, this study suggests viral compartmentalization in the semen microenvironment after an intensification and stimulatory protocol, with evidence of viral evolution. We conclude that the seminal microenvironment was not the source of the blood viral rebound, which further supports
the hypothesis of viral compartmentalization.14
In summary, intensification and stimulatory therapy in this study of patients chronically infected with HIV-1 was able to be safely utilized with relatively minimal adverse side effects.10 Moreover, the therapies did not consistently affect the seminal compartment (ED NOTE from Jules Levin: so they think! The seminal virus appears different so they conclude the seminal virus was not the source of the plasma virus, I'm not quite convinced yet, there are holes in this argument I think), which might be desirable when taking into consideration the possibility of increasing risks of sexual transmission during the course of future viral eradication therapeutic strategies. Nonetheless, further studies are necessary to evaluate the role of the male genital tract, during the course of stimulatory therapy or structured treatment interruptions (STI), as a potential viral reservoir. Understanding, in detail, the pathophysiology of residual HIV-1 disease, including proviral latency, residual replication, and potential sanctuary sites, will be critical in the rational design of studies aimed at sustained
HIV-1 remissions and/or viral eradication in the future.
SEQUENCE DATA
The sequences have been submitted to GenBank, and the accession numbers are AY786499 to AY7864505. Perfectly matched DNA-based sequences, within each patient grouping, were not reiterated in the submission.
REFERENCES
1. Palella FJ, Delaney KM, Moorman AC, et al.: Declining morbidity and mortality among patients with advanced human immunodeficiency virus infection. N Engl J Med 1998;338:853-860.
2. Hammer SM, Squires KE, Hughes MD, et al.: A controlled trial
of two nucleoside analogues plus indinavir in persons with human
immunodeficiency virus infection and CD4 cell counts of 200 per
cubic millimeter or less. AIDS Clinical Trials Group 320 Study
Team. N Engl J Med 1997;337:725-733.
3. Gupta P, Mellors J, Kingsley L, et al.: High viral load in semen of
human immunodeficiency virus type 1-infected men at all stages
of disease and its reduction by therapy with protease and nonnucleoside
reverse transcriptase inhibitors. J Virol 1997;71:
6271-6275.
4. Chun TW, Carruth L, Finzi D, et al.: Quantification of latent tissue
reservoirs and total body viral load in HIV-1 infection. Nature.
1997;387:183-188.
5. Dornadula G, Zhang H, VanUitert B, et al.: Residual HIV-1 RNA
in blood plasma of patients taking suppressive highly active antiretroviral
therapy. J Am Med Assoc 1999;282:1627-1632.
6. Finzi D, Hermankova M, Pierson T, et al.: Identification of a reservoir
for HIV-1 in patients on highly active antiretroviral therapy.
Science 1997;278:1295-1300.
7. Zhang H, Dornadula G, Beumont M, et al.: Human immunodeficiency
virus type 1 in the semen of men receiving highly active
antiretroviral therapy. N Engl J Med 1998;339:1803-1809.
8. Pomerantz RJ: Residual HIV-1 infection during antiretroviral therapy:
The challenge of viral persistence. AIDS 2001;15:1201-1211.
9. Sonza S, Mutimer HP, Oelrichs R, et al.: Monocytes harbour replication-
competent, non-latent HIV-1 in patients on highly active antiretroviral
therapy. AIDS 2001;15:17-22.
10. Kulkosky J, Nunnari G, Otero M, et al.: Intensification and stimulation
therapy for human immunodeficiency virus type 1 reservoirs
in infected persons receiving virally suppressive highly active
antiretroviral therapy. J Infect Dis 2002;186:1403-1411.
11. Lori F, Malykh A, Cara A, et al.: Hydroxyurea as an inhibitor of
human immunodeficiency virus type 1 replication. Science
1994;266:801-804.
12. Zhang L, Chung C, Hu B-S, et al.: Genetic characterization of rebounding
of HIV-1 after cessation of highly active antiretroviral
therapy. J Clin Invest 2000;106:839-845.
13. Chun T-W, Davey RT, Ostrowski M, et al.: Relationship between
pre-existing viral reservoirs and the re-emergency of plasma
viremia after discontinuation of highly active antiretroviral therapy.
Nat Med 2000;6:757-761.
14. Nunnari G, Otero M, Dornadula G, et al.: Residual HIV-1 disease
in seminal cells of HIV-1-infected men on suppressive HAART:
Latency without on-going cellular infections. AIDS 2002;16:
39-45.
15. Kulkosky J, Sullivan J, Xu Y, et al.: Genotypic alteration of
HAART-persistent HIV-1 reservoirs in vivo. Virology 2003;314:
617-629.
|
|
| |
| |
|
|
|