| |
HIV-like virus found in wild chimps
|
| |
| |
Discovery supports theory that human HIV pandemic came from African apes.
Published online: 25 May 2006
Nature.com
Michael Hopkin

SIV has finally been tracked down in wild west African chimps.
credit: Ian C. Gilby
Scientists have spotted the signs of an HIV-like virus in chimpanzees in southern Cameroon, confirming the long-held suspicion that these animals are a natural reservoir for the virus in the wild.
The discovery bolsters the theory that the first people to contract HIV did so through contact with infected blood from wild chimps in the jungle, before eventually spreading the virus to nearby Kinshasa in the Democratic Republic of Congo and onwards from there.
Researchers led by Beatrice Hahn of the University of Alabama at Birmingham travelled to Cameroon to collect droppings from the chimpanzee subspecies Pan troglodytes troglodytes.
The team knew that a few captive chimps of this subspecies have been found carrying a strain of simian immunodeficiency virus (SIV) almost identical to the HIV-1 strain, but it was not clear how these animals came to have the virus. Better evidence that these creatures were responsible for the human AIDS pandemic would come from tracking down a reservoir of the human-like SIV in wild chimps in west Africa. The team's hopes of finding such a reservoir were high: a few years ago they found another version of SIV, one quite different from the human virus, in a different subspecies of wild chimp living in east Africa.
As hoped, analysis of the Cameroon samples revealed the presence of antibodies against human-like SIV and traces of the virus' genetic sequence. On the basis of their samples, the researchers calculate that some 30-35% of chimpanzees are carriers. The team reports the findings in Science1.
The virus does not seem to cause any AIDS-like symptoms in the chimpanzees, says Hahn, as captive infected chimps do not seem to develop immune disease. "Lots of people are trying to find out why," says Paul Sharp, a viral geneticist at the University of Nottingham, UK, who also worked on the study.
Wild hunt
The virus probably got into humans as a result of bushmeat hunting, Hahn suggests. "The most likely route, based on the biology of these viruses, is human exposure to infectious chimp blood or body fluids during hunting and butchering," she says.
The genetics of the wild chimp SIV are very close to the human virus. This suggests that humans contracted the virus directly from chimps, rather than both humans and chimps contracting it from monkeys, as some experts had previously suggested.
It is unclear exactly how the virus arose in the chimps themselves, but it is probably derived from two viruses carried by monkey species on which the chimps prey, suggests Sharp.
River commute
Analysis of the path of the human pandemic has pinpointed Kinshasa as the epicentre of the outbreak. The first HIV-positive human blood was obtained here in 1959. The virus was almost certainly carried here by infected humans, says Sharp: "Chimps don't walk the streets of Kinshasa."
The theory is supported by the fact that rivers, the primary transport routes through the dense West African jungle, provide an easy means of travelling from southern Cameroon to Kinshasa. Once there, the urban environment would have been far more conducive to the transmission of the virus between people. But it will be a difficult story to verify with certainty. "We're talking about something thought to have happened more than 75 years ago," he says.
It seems to be largely a matter of chance that this strain became the one to terrorize the globe, says Sharp. HIV-2, thought to have come from sooty mangabey monkeys, also causes AIDS in humans, but this virus is confined almost entirely to West Africa.
Chimpanzee Reservoirs of Pandemic and Nonpandemic HIV-1
"......The findings presented here, together with prior studies, provide for the first time a clear picture of the origin of HIV-1 and the seeds of the AIDS pandemic. SIVcpz, the progenitor of HIV-1, arose as a recombinant of ancestors of SIV lineages presently infecting red-capped mangabeys and Cercopithecus monkeys in west-central Africa..... HIV-1 groups M, N and O each resulted from independent crossspecies transmissions of SIVcpzPtt from P. t. troglodytes to man early in the 20th century..... it is quite possible that still other SIVcpz lineages exist that could pose risks for human infection and prove problematic for HIV diagnostics and vaccines...."
Science Express May 25, 2006
Brandon F. Keele,1 Fran Van Heuverswyn,2 Yingying Li,1 Elizabeth Bailes,3 Jun Takehisa,1 Mario L. Santiago,1* Frederic Bibollet-Ruche,1 Yalu Chen,1 Louise V. Wain,3 Florian Liegeois,2 Severin Loul,4 Eitel Mpoudi Ngole,4 Yanga Bienvenue,4 Eric Delaporte,2 John F. Y. Brookfield,3 Paul M. Sharp,3 George M. Shaw,1,5 Martine Peeters,2 Beatrice H. Hahn1
1Departments of Medicine and Microbiology, University of Alabama at Birmingham, Birmingham, AL, USA. 2Laboratoire Retrovirus, UMR145, Institut de Recherche pour le Developpement (IRD) and Department of International Health, University of Montpellier I, 911 Avenue Agropolis, BP 64501, 34394 Montpellier Cedex 5, France. 3Institute of Genetics, University of Nottingham, Queens Medical Centre, Nottingham, NG7 2UH, UK. 4Projet Prevention du Sida au Cameroun (PRESICA), BP1857, Yaounde, Cameroun. 5Howard Hughes Medical Institute, 720 South 20th Street, KAUL 816, Birmingham, AL 35294, USA.
Human immunodeficiency virus type 1 (HIV-1), the cause of human AIDS, represents a zoonotic infection of staggering proportions and social impact. Yet, uncertainty persists regarding its natural reservoir. The virus most closely related to HIV-1 is a simian immunodeficiency virus (SIV) thus far identified only in captive members of the chimpanzee subspecies Pan troglodytes troglodytes. Here, we report the detection of SIVcpz antibodies and nucleic acids in fecal samples from wild-living P. t. troglodytes apes in southern Cameroon where prevalence rates in some communities reached 29 to 35%. By sequence analysis of endemic SIVcpz strains, we could
trace the origins of pandemic (group M) and nonpandemic (group N) HIV-1 to distinct, geographically isolated chimpanzee communities. These findings establish P. t. troglodytes as the natural reservoir of HIV-1.
Since the first detection of an HIV-1 related lentivirus in chimpanzees (1, 2), this species has been suspected as the source of the human AIDS pandemic. However, a crucial missing link in the chain of evidence implicating SIVcpz in
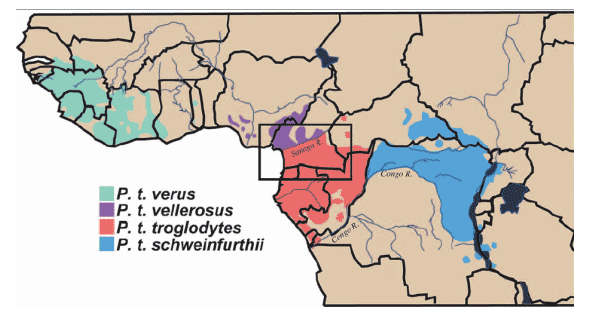
the origin of HIV-1/AIDS has been the absence of a recognizable virus reservoir in wild-living apes. Chimpanzees (Pan troglodytes) are classified into four subspecies on the basis of differences in mitochondrial DNA sequence (3): P. t.
verus in west Africa; P. t. vellerosus in Nigeria and northern Cameroon; P. t. troglodytes in southern Cameroon, Gabon, and the Republic of Congo; and P. t. schweinfurthii in the Democratic Republic of Congo and countries to the east (Fig. 1). Two of these subspecies, P. t. troglodytes and P. t. schweinfurthii, are known to harbor SIVcpz, and their viruses form divergent subspecies-specific phylogenetic lineages (4). HIV-1 is most closely related to SIVcpzPtt (5), but this virus has been detected only rarely, and then only in captive apes (1, 5-7). There is no counterpart of SIVcpzPts that is known to infect humans (4, 8-10).
Wild-living chimpanzees are reclusive, highly endangered, and live in remote jungle areas. To study chimpanzees in their natural habitat, we developed methods to detect SIVcpz specific antibodies and nucleic acids in fecal samples
collected from the forest floor (9-11). In addition, we developed genotyping approaches to amplify host mitochondrial and genomic markers (polymorphic
microsatellite loci) from these same specimens for species, gender and individual identification (11, 12). These methods were validated in captive and habituated apes of known infection status (13). Here, we employed these non-invasive
approaches to conduct the first molecular epidemiological field study of SIVcpz in wild-living, non-habituated chimpanzees in west central Africa.
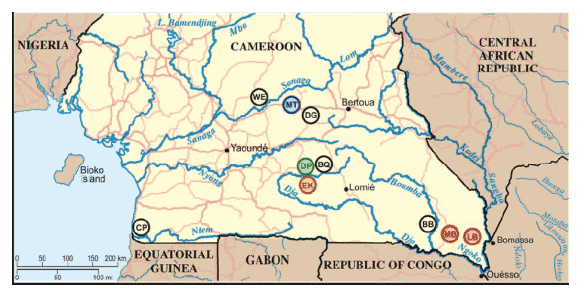
Cameroon is home to two chimpanzee subspecies, P. t. vellerosus in the north and P. t. troglodytes in the south, with the Sanaga River forming the boundary between their ranges (Fig. 1). In the present study, we collected 599 fecal
specimens at 10 forest sites throughout the southern part of Cameroon (Fig. 1). All field sites, except one (WE), were in the range of the P. t. troglodytes subspecies. To establish the species and subspecies origin of each sample, a 498 bp mitochondrial DNA (D loop) fragment was amplified from fecal DNA and subjected to phylogenetic analysis (13).
Eighty-six specimens were degraded, while 67 samples contained gorilla mtDNA sequences (table S1). The remaining 446 samples were of chimpanzee origin, 423 from P. t. troglodytes and 23 from P. t. vellerosus. These comprised 82 unique mtDNA haplotypes (fig. S1 and table S2). Consistent with the recognized ranges of the two subspecies, all 23 P. t. vellerosus specimens were collected
north of the Sanaga River, whereas 421 of 423 P. t. troglodytes samples were collected to the south (table S1).
All mtDNA positive fecal samples were tested for virus specific antibodies using a sensitive immunoblot assay specifically developed for surveys at remote field sites (13). This analysis identified 34 specimens, all from P. t. troglodytes apes, that contained antibodies reactive with HIV-1 antigens (Fig. 2). Twelve samples exhibited a strong and broadly cross-reactive Western blot profile that was virtually indistinguishable from the HIV-1 positive human plasma control. Eighteen additional samples reacted with both the HIV-1 envelope (gp160) and major core (p24) proteins, thus also meeting formal criteria for HIV-1/SIVcpz antibody positivity. Four samples (EK502, EK506, MB245, MB248) reacted only faintly with a single HIV-1 protein (p24) and were classified as indeterminant. None of 23 P. t. vellerosus or 67 gorilla specimens exhibited detectable Western blot reactivity to any HIV-1 protein (table S1).
To corroborate the fecal antibody results, RNA was extracted from all immunoblot reactive samples and subjected to RT-PCR amplification using consensus env and pol primers. In addition, fecal DNA was used to amplify
polymorphic microsatellite loci to identify and distinguish individual apes, and to amplify a portion of the amelogenin gene for gender determination (13). These analyses revealed that the 34 immunoblot reactive samples represented 16
different P. t. troglodytes apes (7 males and 9 females). Each of these apes had detectable virion RNA in one or more fecal samples (table S3). SIVcpz env (~390 bp) and/or pol (~890 bp) sequences were amplified from 31 of 34 (91%)
immunoblot reactive samples, including all four specimens with indeterminant Western blot reactivity (Fig. 3 and table S3). These data, together with previous findings for SIVcpzPts-infected apes (10), indicate that fecal antibody reactivity to a single HIV-1 Gag protein is indicative of SIVcpzPtt infection (14).
The prevalence of SIVcpzPtt infection in wild chimpanzee communities was estimated for each of the 10 field sites (table S1). For the DP, EK, MB, BB and LB communities, this was done based on the proportion of infected individuals
as determined by microsatellite analyses, taking into consideration assay sensitivities and specimen degradation (tables S1 and S4). For the remaining sites, prevalence rates were estimated based on the proportion of antibody and/or SIVcpz vRNA positive fecal samples, while also adjusting for repeat sampling (13). The results indicated widespread but notably uneven SIVcpzPtt infection of wild-living P. t. troglodytes apes, with prevalence rates ranging from 23% to 35% in the LB, EK and MB communities, 4% to 5% in the DP and MT communities, and absence of infection in the WE, DG, BQ, BB and CP communities.
To determine the evolutionary relationships of the 16 new SIVcpzPtt viruses to each other and to previously characterized SIVcpz and HIV-1 strains, pol and env sequences were subjected to phylogenetic analyses. All of the newly identified SIVcpz strains were found to fall within the radiation of SIVcpzPtt strains from captive P. t. troglodytes apes, which also includes HIV-1 groups M and N but not group O or SIVcpzPts (Fig. 3). Interestingly, the new P. t.
troglodytes viruses exhibited significant phylogeographic clustering: SIVcpz sequences from the EK, DP, MT and MB/LB collection sites formed well-separated clades corresponding to their field site of origin. One of these clades
included closely related SIVcpz strains (EK519, EK516, EK502, EK505), likely reflecting recent virus transmission within that community. The remaining clades were each comprised of more divergent but still monophyletic SIVcpz strains (Fig. 3). Thus, chimpanzee populations separated by long distances or major geographical barriers such as rivers (Fig. 1) harbored distinct SIVcpz lineages (e.g., EK, DP, and MT), while neighboring communities not separated by such
barriers harbored viruses that were phylogenetically interspersed (e.g., MB and LB).
The phylogeographic clustering of the newly identified SIVcpzPtt strains allowed us to trace the origins of present day human AIDS viruses to distinct chimpanzee
communities. In subgenomic pol and env regions, SIVcpzPtt strains from the MB/LB and EK sites were much more closely related to HIV-1 groups M and N, respectively, than were any previously identified SIVcpz strains (Fig. 3). Fulllength genome analysis of four of the 16 new viruses confirmed and extended these findings, revealing strong statistical support for the clustering of HIV-1 groups M and N with the MB/LB and EK lineages of SIVcpzPtt, respectively
(Fig. 4). Moreover, inclusion of the new viruses reduced the lengths of the branches marking the cross-species transmission events for all genomic regions by almost half (see arrows in Fig. 4). Given these short branch lengths, it is
highly unlikely that other SIVcpzPtt strains exist that are significantly more closely related to HIV-1 groups M and N than are the viruses from the MB/LB and EK communities. Indeed, expanded field studies in southern Cameroon by our
group have identified additional SIVcpzPtt strains, including nine from the MB/LB area, whose sequences support this conclusion and corroborate the phylogenetic relationships shown in Figs. 3 and 4 (15). Thus, an extensive set of molecular epidemiological data all points to chimpanzees in southeastern and southcentral Cameroon as the sources of HIV-1 groups M and N, respectively.
The findings presented here, together with prior studies, provide for the first time a clear picture of the origin of HIV-1 and the seeds of the AIDS pandemic. SIVcpz, the progenitor of HIV-1, arose as a recombinant of ancestors of SIV lineages presently infecting red-capped mangabeys and Cercopithecus monkeys in west-central Africa (16). Chimpanzees acquired this recombinant virus, or its progenitors, by cross-species transmission some time after the split of P. t. verus and P. t. vellerosus from the other subspecies (fig. S1), but possibly before the divergence of P. t. schweinfurthii from P. t. troglodytes (4). This explains the absence of SIVcpz infection in present day P. t. verus and P. t. vellerosus apes, the presence of SIVcpz infection in P. t. troglodytes and P. t. schweinfurthii apes, and the phylogenetic separation of SIVcpzPtt from SIVcpzPts viruses (4, 7, 9, 15). HIV-1 groups M, N and O each resulted from independent crossspecies transmissions of SIVcpzPtt from P. t. troglodytes to man early in the 20th century (17-19). We show here that the SIVcpzPtt strain that gave rise to HIV-1 group M belonged to a viral lineage that persists today in P. t. troglodytes apes in southeastern Cameroon. That virus was likely transmitted locally. From there it appears to have made its way via the Sangha River (or other tributaries) south to the Congo River and on to Kinshasa where the group M pandemic was likely spawned (20). HIV-1 group N, which has been identified in only a small number of AIDS patients from Cameroon (21, 22), derived from a second SIVcpzPtt lineage in southcentral Cameroon and remained geographically more restricted. The source of HIV-1 group O remains unknown but will likely yield to further study of wild ape populations not yet sampled. Given the extensive genetic diversity and phylogeographic clustering of SIVcpz now recognized, and the vast areas of west central Africa not yet sampled (Fig. 1), it is quite possible that still other SIVcpz lineages exist that could pose risks for human infection and prove problematic for HIV diagnostics and vaccines. The present report describes molecular tools and non-invasive strategies that can be used to explore these possibilities as well as the molecular ecology of pathogens in endangered species more generally.
References and Notes
1. M. Peeters et al., AIDS 3, 625 (1989).
2. T. Huet et al., Nature 345, 356 (1990).
3. C. P. Groves, in Mammalian Species of the World: A Taxonomic and Geographic Reference, D. E. Wilson, D. M. Reader, Eds. (Smithsonian Institution Press, Washington, DC, ed. 2, 1993), pp. 243-277.
4. P. M. Sharp, G. M. Shaw, B. H. Hahn, J. Virol. 79, 3891 (2005).
5. F. Gao et al., Nature 397, 436 (1999).
6. S. Corbet et al., J. Virol. 74, 529 (2000).
7. E. Nerrienet et al., J. Virol. 79, 1312 (2005).
8. M. M. Vanden Haesevelde et al., Virology 221, 346 (1996).
9. M. L. Santiago et al., Science 295, 465 (2002).
10. M. Worobey et al., Nature 428, 820 (2004).
11. M. L. Santiago et al., J. Virol. 77, 7545 (2003).
12. M. L. Santiago et al., J. Virol. 79, 12515 (2005).
13. Materials and methods are available as supporting material on Science Online.
14. In contrast to plasma samples from uninfected humans which exhibit false positive Western blot reactivity to HIV-1 p24 in as many as 10 - 15% of individuals (www.fda.gov/cber/products/hiv1cam052898.htm), we have found no such nonspecific cross-reactivity of chimpanzee immunoglobulin extracted by the RNAlater method from over 2,000 fecal specimens.
15. F. Van Heuverswyn et al., 13th Conference on Retroviruses and Opportunistic Infections, Abstr. 132, available at
http://www.retroconference.org/2006/Abstracts/26513.htm (2006).
16. E. Bailes et al., Science 300, 1713 (2003).
17. B. H. Hahn, G. M. Shaw, K. M. De Cock, P. M. Sharp, Science 287, 607 (2000).
18. B. T. K. Korber et al., Science 288, 1789 (2000).
19. P. M. Sharp et al., Philos. Trans. R. Soc. London Ser. B 356, 867 (2001).
20. N. Vidal et al., J. Virol. 74, 10498 (2000).
21. F. Simon et al., Nature Medicine 4, 1032 (1998).
22. J. Yamaguchi et al., AIDS Res. Hum. Retroviruses 22, 83 (2006).
23. We thank the Cameroonian Ministries of Health, Environment and Forestry, and Research for permission to perform this study; J. Dupain, C. Neel, and M. Epanda (Projet Grands Singes); L. Usongo, D. Dontego, F. Espedi, and B. Tshikangwa (World Wildlife Fund); and G. Etoga and D. M'bohand (Ministry of Environment and Forestry) for assistance in the field. This work was supported in part by the NIH (R01 AI50529, R01 AI58715, P30 AI 27767), the Bristol Myers Freedom to Discover Program, the Institut de Recherche pour le Developpement, and the Howard Hughes Medical Institute. New SIVcpzPtt sequences are available at GenBank under accession numbers DQ370366-DQ370419 and DQ373063-DQ373066; chimpanzee mtDNA sequences are available under DQ367532-DQ367613 and DQ370307-DQ370365.
|
|
| |
| |
|
|
|