| |
Oral HIV-exposure elicits mucosal HIV-neutralizing antibodies in uninfected men who have sex with men
|
| |
| |
[BASIC SCIENCE: CONCISE COMMUNICATION]
AIDS:Volume 23(3)28 January 2009p 329-333
Hasselrot, Klaraa; Saberg, Parb; Hirbod, Tahaa; Soderlund, Johana; Ehnlund, Mariethea; Bratt, Goranb; Sandstrom, Ericb; Broliden, Kristinaa
aDepartment of Medicine, Solna, Unit of Infectious Diseases, Center for Molecular Medicine, Karolinska Institutet, Karolinska University Hospital, Sweden
bGay Men's Health Clinic (Venhalsan), South Hospital, Stockholm, Sweden._
Abstract
Objective: To determine whether oral sexual exposure to HIV-1 (HIV) results in HIV-neutralizing activity in saliva of uninfected men who have sex with infected men?
Design: Saliva samples were collected from HIV IgG seronegative men (n = 25) whose male partners were HIV infected and from low-risk healthy controls (n = 22) and analyzed for HIV-neutralizing capacity.
Methods: The presence of neutralizing activity in saliva was tested in a peripheral blood mononuclear cell-based assay using primary HIV isolates. Self-reporting questionnaires described the individuals' sexual behaviors and routes of possible HIV exposure.
Results: Of 25 exposed, uninfected individuals (EUI), 21 reported receptive unprotected oral intercourse, whereas three of the 25 reported unprotected anal receptive intercourse. Whole saliva from both EUI and low-risk healthy controls contained HIV-neutralizing activity. However, a significant difference was seen when analyzing the salivary IgA1 fraction: 13 of 25 EUI neutralized HIV, whereas none of the 22 controls had this capacity. The neutralizing capacity of the EUI males persisted during 2 years of follow-up.
Conclusion: Unprotected oral sex evokes a salivary IgA1-mediated HIV-neutralizing response that persists over time during continuous exposure in uninfected male partners of infected men.
Introduction
The oral transmission of HIV-1 (HIV) from infected mothers to nursing infants is an unquestioned route of infection mainly in countries with predominantly low-income populations. Furthermore, the body of evidence regarding oral transmission in adult human populations, via oral sexual intercourse, is growing. Although the mechanisms of transmitting HIV orally need better definition, it is clear that tonsilar lymphoid tissues are susceptible to infection [1-3]. We know that the virus is present in both seminal and preseminal fluid [4], and several case studies have documented exclusively oral sex with an infected individual as the probable cause of HIV transmission [5-8]. However, studies of the corresponding mucosal immune responses to HIV exposure during oral sex are rare, especially in men who have sex with men. Yet, this issue is of considerable importance for the understanding of viral transmission and for the development of new prophylactic measures.
Research on HIV-exposed uninfected individuals (EUI) has defined several immune responses and genetic factors that are associated with protection against mucosal HIV transmission [9]. Indeed, an association of genital mucosal HIV-neutralizing IgA1 antibodies and HIV nonacquisition was recently found in a prospective study on HIV-exposed women [10]. To further this investigation, we examined a previously unpublished cohort of HIV-serodiscordant male couples to determine whether oral exposure to HIV elicits neutralizing salivary activity and, if so, whether this response persists over time.
Methods
Study cohort
EUI were recruited via their HIV-positive partners regularly attending the Gay Men's Health Clinic in Stockholm, Sweden. Inclusion criteria were a HIV-negative man engaged in at least a 6-month relationship with a male HIV-positive partner. The 25 study individuals were followed regularly for 2 years. Each study individual received a questionnaire regarding demographical data and sexual behaviors. At all visits, each participant tested negative for HIV, chlamydia and gonorrhea. Research nurses collected all samples during the evening from EUI given directions not to eat or drink for the preceding 2 h. Whole saliva was collected (3-5 ml/visit) in 50 ml vials, divided into aliquots and frozen at -80°C within 2 h. Control individuals (n = 22), all HIV-negative healthy men, were recruited by advertisement at a blood donor clinic. The HIV-positive partners were not personally involved in the study; however, their clinical data were available for the study. The study was approved by the ethical committee at Karolinska Institutet. All participants gave their informed consent.
IgA1-purification and IgA1 quantification
Samples of saliva were purified with jacalin-agarose (Vector Labs, Burlingame, California, USA); 266 _l of the saliva samples were diluted with 534 _l PBS pH 7.4 (1: 3 dilution resulting in a total amount of 800 _l) were added to 200 _l jacalin/agarose beads and mixed for 2 h at +4°C followed by centrifugation. The jacalin-agarose beads were thoroughly washed with PBS, pH 7.4, after which the bound IgA1 was eluted overnight at +4°C by adding 1ml 0.8 mol/l D-galactose, pH 7.4. The supernatant was subsequently collected; all fractions were stored at -80°C. An inhouse enzyme-linked immunosorbent assay (ELISA) was used for total IgA1 quantification [11].
HIV-1 neutralization assay
Viruses used in the HIV-neutralization assay were obtained from the NIH Aids Research and Reference Reagent Program (USA) and included BZ-167 (X4R5-virus, Brazil) and 92/US/727 (R5-virus, USA). TCID50 values were determined for each isolate, and three virus dilutions (ranging from TCID50 _ 10-30) were tested at each time to compensate for peripheral blood mononuclear cells (PBMC) donor variability. The optimal dilution for whole saliva was found to be 1: 10, whereas the IgA1 fractions were used without further dilution (through the purification process they were already diluted 1: 4). The neutralization assays were performed according to a predefined protocol [12]. In brief, duplicate wells of each virus dilution and each sample fraction were incubated for 1 h at 37°C, followed by the addition of 1 _ 105 PHA-stimulated PBMC, pooled from two individual donors. The cells were washed at days 1, 4, and 6 of incubation at 37°C; supernatant from each well was collected at day 6 and analyzed in a p24 antigen ELISA (Vironostika HIV-1 Antigen kit; Biomerieux, Boxtel, The Netherlands). Neutralization capacity was defined as a more than 67% reduction in the supernatant as compared with p24-antigen content in the presence of previously standardized low-risk healthy control samples.
Results
Saliva samples of exposed uninfected individuals can neutralize HIV
Saliva from a total of 25 EUI and 22 low-risk healthy controls were tested for HIV-neutralizing activity. In the whole saliva fractions, 15 of the EUI and six of the controls had HIV-neutralizing activity. However, when IgA1 antibodies were purified from the samples, a striking difference emerged: 13 of 25 EUI effectively neutralized the BZ-167 HIV isolate, as compared with none of 22 of the controls (Fischer's exact test: P = 0.001). The individuals whose samples neutralized BZ-167 were also tested against an additional HIV-primary isolate (92/US/727), and 11 of the 13 samples exhibited the same neutralizing ability. The neutralization capacity of IgA1 is likely to be a significant contributor in the whole-saliva samples; firstly - the magnitude of neutralization by purified IgA1 was similar to that of whole saliva, secondly - when testing the 'non-IgA1-fraction' (whole sample but IgA1 depleted) from the 13 EUI who neutralized by IgA1, eight individuals could not neutralize (data not shown).
To question whether the neutralizing capacity persisted in EUI during ongoing relationships, we tested IgA1 from the last salivary sampling of each individual, a median of 23 months after enrollment. The ability to neutralize was largely unchanged; one of 13 EUI lost this capacity, and one additional EUI was now able to neutralize the BZ-167 isolate. At this later time point, all EUI remained HIV-uninfected and were within the same relationship.
Quantity of salivary IgA1
The quantities of total IgA1 in saliva were significantly higher among the EUI (median 465 μg/ml, range 100-1100 μg/ml) than in the control individuals (median 284 μg/ml, 50-950 μg/ml) (P = 0.019: Mann-Whitney).
Estimate of sexual exposure to HIV
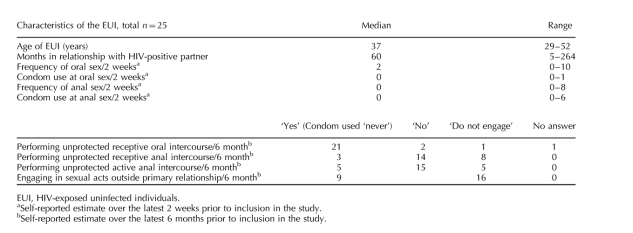
To determine the extent of HIV exposure, all EUI described their sexual habits during the 2 weeks immediately before filling out the investigators' questionnaire. Additionally, the frequency of sexual acts and protection strategies during the latest 6 months of each relationship were summarized (Table 1). The responses indicated that unprotected receptive oral intercourse had been performed with the HIV-positive partner by 21 of 25 EUI during the 6 months before questioning. The corresponding figure for unprotected receptive anal intercourse was three of 25. By the time of inclusion in the study, 22 of 25 HIV-positive partners were receiving antiretroviral treatment (ART) and the remaining three were untreated with low virus levels and normal CD4 cell counts. The highest viral loads in the infected partners during the relationship had occurred before their inclusion in the study (data not shown), and they had known about their HIV serostatus for 1 to 18 years (median 11 years). Only two of the partners had been on ART with low viremia during the whole relation. Thus, 23 of 25 EUI were likely exposed to HIV due to the HIV-infected partner's: initial unawareness of HIV-serostatus, lack of treatment or ART failure. Possible HIV exposure continued throughout the 2-year study for some individuals due to interruption of ART or treatment failures in combination with unprotected sexual intercourse.
Table 1. Characteristics of the study individuals and estimated sexual HIV exposures.
Discussion
Here we report for the first time that exposure to HIV, as a consequence of oral intercourse, is sufficient to produce an IgA1-mediated HIV-neutralizing response in the oral mucosa of uninfected men with a male partner. Previously, HIV-exposed, uninfected women have been studied extensively with regard to mucosal antiviral immune responses; however, the focus in those studies was on genital interactions in a heterosexual setting [9,10]. In the present study, a group of 25 uninfected male partners of HIV-infected men were requested to donate mucosal samples and undergo thorough self-documentation of sexual behaviors, an acknowledged and reliable method of exploring these issues [13]. The clear pattern that emerged was that oral sex was considered to be of low risk and was at almost all times performed without protection from seminal emissions, whereas anal intercourse was considered as a high-risk act and was therefore avoided or performed with condoms. The majority of the HIV-infected partners experienced periods of uncontrolled viremia and the corresponding EUI were thus potentially exposed to HIV during unprotected sexual intercourse. The significant result of the present study was that 13 of 25 EUI had HIV-neutralizing IgA1 in their salivary samples, which could not be found in any of the samples from 22 males in the control group. Furthermore, 12 of the 13 EUI retained their capacity to neutralize the virus for 2 years after the study began. At this final time point, one additional EUI had acquired salivary neutralizing IgA1 activity, despite an earlier negative report.
Although we did not find any correlation between the ability to neutralize HIV and the level of self-reported unprotected sexual intercourse, we did see a trend when correlating the peak viral load in the corresponding partner during the relation with the neutralization capacity of the EUI (median 199 300copies/ml in partners of neutralizing EUI, median 10 800copies/ml in partners of nonneutralizing EUI, P = 0.10, data not shown). This could suggest that the EUI's ability to neutralize HIV is determined by the level of viral exposure.
The natural anti-HIV capacity of whole saliva is well known and was confirmed in some of the EUI and control individuals. This biological activity is most likely mediated by chemokines and other antimicrobial molecules [11]. The neutralizing capacity of whole saliva is not likely to be mediated by non-HIV-specific IgG, as purified IgG from salivary samples (both EUI and controls) were not able to neutralize HIV (data not shown). The anti-HIV activity of the salivary IgA1 from the EUI could theoretically be mediated by contaminating soluble molecules, but this seems unlikely, as no anti-HIV activity was observed in salivary IgA1 from the low-risk controls. Seminal remnants from the HIV-infected partners could also theoretically contribute to the neutralizing activity of the saliva, but this possibility was rejected by the results of testing that proved negative for prostate-specific antigen in all cases (data not shown). Unfortunately, no reliable HIV-binding assay for IgA antibodies is available [14], and the epitope specificity of IgA1 antibodies has not been evaluated. However, possible IgA specificity for cellular receptors such as CCR5 [15] or for nonenvelope HIV proteins [16] or carbohydrates cannot be excluded. Such induced or natural antibodies in mucosal secretions could provide a first-line host defense against HIV infection.
In this report, the initial use of a viral isolate, BZ-167, that is reported to be moderately neutralization sensitive and careful evaluation of alternative assay conditions provided robust and reproducible data. The alternative-assay conditions included an appraisal of PBMC donor variability and of a second HIV-primary isolate [17-19]. The isolates used in the present assay were of the R5 and R5X4 dualtropic phenotype, thereby representing viral phenotypes to which the individuals of this study were likely to be exposed. As salivary antibodies and innate molecules are the first and most superficial natural barriers to halt the transmission of virus through the oral mucosal, they must have the capacity to inhibit multiple viral phenotypes. The selective transmission of R5 virus, which under certain circumstances leads to a systemic infection, may be determined by the properties of the foregoing barriers and/or those that a virus subsequently encounters in the epithelium and target cells.
Oral receptive sex, a behavior commonly performed by both partners in male/male couples, has previously been viewed as a possible means of HIV transmission [20,21]. Although a low transmission rate per act is the likely outcome, the risk cannot be discounted if the frequency of these acts is high. Presumably, oral sex practices occur repetitively and probably increasingly due to the long-held conviction that they constitute 'safe sex' [22]. Prospective studies in similar cohorts, as well as retrospective studies using the corresponding partner's own viral isolate, would complement our findings and further determine the contribution of this humoral immune response within the mucosa to protecting recipients of oral sex from HIV infection.
References
1. Baba TW, Trichel AM, An L, Liska V, Martin LN, Murphey-Corb M, et al. Infection and AIDS in adult macaques after nontraumatic oral exposure to cell-free SIV. Science 1996; 272:1486-1489.
2. Maher DM, Zhang ZQ, Schacker TW, Southern PJ. Ex vivo modeling of oral HIV transmission in human palatine tonsil. J Histochem Cytochem 2005; 53:631-642.
3. Stahl-Hennig C, Steinman RM, Tenner-Racz K, Pope M, Stolte N, Matz-Rensing K, et al. Rapid infection of oral mucosal-associated lymphoid tissue with simian immunodeficiency virus. Science 1999; 285:1261-1265.
4. Pudney J, Oneta M, Mayer K, Seage G 3rd, Anderson D. Preejaculatory fluid as potential vector for sexual transmission of HIV-1. Lancet 1992; 340:1470.
5. DeGruttola V, Mayer KH. Human immunodeficiency virus and oral intercourse. Ann Intern Med 1987; 107:428-429.
6. Goldberg DJ, Green ST, Kennedy DH, Emslie JA, Black JD. HIV and orogenital transmission. Lancet 1988; 2:1363.
7. Keet IP, Albrecht van Lent N, Sandfort TG, Coutinho RA, van Griensven GJ. Orogenital sex and the transmission of HIV among homosexual men. AIDS 1992; 6:223-226.
8. Schacker T, Collier AC, Hughes J, Shea T, Corey L. Clinical and epidemiologic features of primary HIV infection. Ann Intern Med 1996; 125:257-264.
9. Kulkarni PS, Butera ST, Duerr AC. Resistance to HIV-1 infection: lessons learned from studies of highly exposed persistently seronegative (HEPS) individuals. AIDS Rev 2003; 5:87-103.
10. Hirbod T, Kaul R, Reichard C, Kimani J, Ngugi E, Bwayo JJ, et al. HIV-neutralizing immunoglobulin A and HIV-specific proliferation are independently associated with reduced HIV acquisition in Kenyan sex workers. AIDS 2008; 22:727-735.
11. Hirbod T, Reichard C, Hasselrot K, Soderlund J, Kimani J, Bwayo JJ, et al. HIV-1 neutralizing activity is correlated with increased levels of chemokines in saliva of HIV-1-exposed uninfected individuals. Curr HIV Res 2008; 6:28-33.
12. Devito C, Hinkula J, Kaul R, Kimani J, Kiama P, Lopalco L, et al. Cross-clade HIV-1-specific neutralizing IgA in mucosal and systemic compartments of HIV-1-exposed, persistently seronegative subjects. J Acquir Immune Defic Syndr 2002; 30:413-420.
13. Carballo-Dieguez A, Remien R, Dolezal, C, Wagner G. Reliability of sexual behaviour self-reports in male couples of discordant HIV status. Journal of Sex Research 1999; http://findarticles.com/p/articles/mi_m2372/is_2_36/ai_54882517 Accessed: 10 December 2008.
14. Wright PF, Kozlowski PA, Rybczyk GK, Goepfert P, Staats HF, VanCott TC, et al. Detection of mucosal antibodies in HIV type 1-infected individuals. AIDS Res Hum Retroviruses 2002; 18:1291-1300.
15. Bouhlal H, Hocini H, Quillent-Gregoire C, Donkova V, Rose S, Amara A, et al. Antibodies to C-C chemokine receptor 5 in normal human IgG block infection of macrophages and lymphocytes with primary R5-tropic strains of HIV-1. J Immunol 2001; 166:7606-7611.
16. Wright A, Yan H, Lamm ME, Huang YT. Immunoglobulin A antibodies against internal HIV-1 proteins neutralize HIV-1 replication inside epithelial cells. Virology 2006; 356:165-170.
17. Mascola JR, D'Souza P, Gilbert P, Hahn BH, Haigwood NL, Morris L, et al. Recommendations for the design and use of standard virus panels to assess neutralizing antibody responses elicited by candidate human immunodeficiency virus type 1 vaccines. J Virol 2005; 79:10103-10107.
18. Li M, Gao F, Mascola JR, Stamatatos L, Polonis VR, Koutsoukos M, et al. Human immunodeficiency virus type 1 env clones from acute and early subtype B infections for standardized assessments of vaccine-elicited neutralizing antibodies. J Virol 2005; 79:10108-10125.
19. Polonis VR, Brown BK, Borges AR, Zolla-Pazner S, Dimitrov DS, Zhang MY, et al. Recent advances in the characrterization of HIV-1 neutralization assays for standardized evaluation of the antibody response to infection and vaccination. Virology 2008; 375:315-320.
20. Page-Shafer K, Veugelers PJ, Moss AR, Strathdee S, Kaldor JM, van Griensven GJ. Sexual risk behavior and risk factors for HIV-1 seroconversion in homosexual men participating in the Tricontinental Seroconverter Study, 1982-1994. Am J Epidemiol 1997; 146:531-542.
21. Bratt GA, Berglund T, Glantzberg BL, Albert J, Sandstrom E. Two cases of oral-to-genital HIV-1 transmission. Int J STD AIDS 1997; 8:522-525.
22. Rothenberg RB, Scarlett M, del Rio C, Reznik D, O'Daniels C. Oral transmission of HIV. AIDS 1998; 12:2095-2105.
|
|
| |
| |
|
|
|