| |
Detectable Viral Load Aggravates Immunosenescence Features of CD8 T-Cell Subsets in Vertically HIV-Infected Children
|
| |
| |
Download the PDF here
JAIDS Journal of Acquired Immune Deficiency Syndromes: 15 August 2012
"Our findings provide evidences of premature immunosenescence in HIV-infected children, notably in viremic children, and reveal the importance of maintaining the HIV-infected children with undetectable viral load.....
"These results indicate that as described in adults,7 HIV infection induces a premature ageing of the effector CD8 pool.
"we emphasize the importance of maintaining undetectable viral load in HIV-infected children to avoid a premature ageing and impairment of the CD8-mediated immune responses."
"Summing up, our results clearly show that HIV infection in children produces a premature ageing of the CD8 T-cell population, notably in children with higher values of viral load. The continuous immune activation of these cells is likely responsible for the appearance of senescence CD8 T-cell markers and the increased rates of apoptosis observed in HIV-infected subjects."
"These results indicate that as described in adults,7 HIV infection induces a premature ageing of the effector CD8 pool. The presence of senescence markers was increased in aviremic children when compared with healthy children, as consequence to the HIV exposition from birth. However, the frequency of all CD8 subsets studied is preserved in this aviremic group and level of senescence was significantly lower than in children with uncontrolled viral load. These data clearly indicate that the control of viral load can reduce dramatically the premature ageing in the CD8 T-cell population."
"persistent antigen exposure to HIV have been associated to hyperactivation of immune system, degradation of immune responses, and AIDS progression. ....It is well known that persistent exposure to HIV-1 antigens accelerates the aging of the immune system in HIV-infected individuals.6,7 Such aging process, similarly to what occurs in the elderly non-HIV-infected people......It is unknown if these HIV-infected children, who are in contact with the HIV since their birth and who suffer from a persistent chronic activation of their immune system from birth, could suffer from a premature "aging", and how it could affect the immune responses."
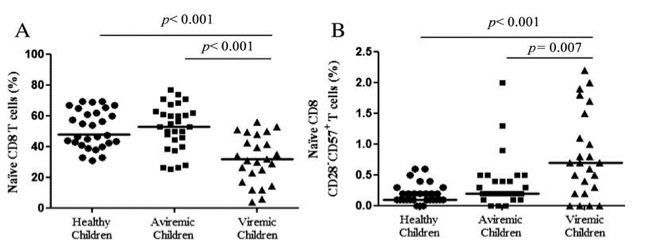
"Naive CD8 T cells of viremic group had an increased frequency of CD28-CD57+ cells in comparison to healthy children (P < 0.001), which were associated to a replicative senescence phenotype. Furthermore, the phenotype of CD8-naive subset in this viremic group is also more senescent (P = 0.007) and proliferative (P = 0.007) than in the aviremic group (Figs. 1B, C). In summary, in aviremic children, the frequency, proliferation, and senescence of naive CD8 T cells are not modified in comparison to healthy children. However, viremic group shows significant alterations in these parameters......in healthy children both naive CD4 and CD8 T cells decrease with age, but percentages of naive CD8 T cells in HIV-infected children are altered and the frequency does not depend on the age."
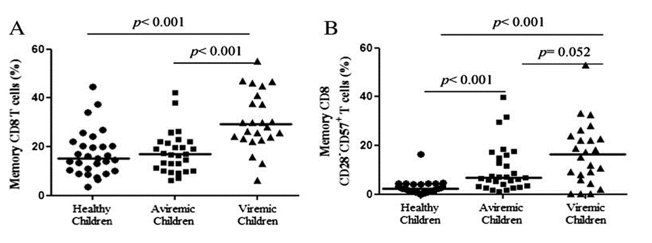
"Memory T-cell subset showed a higher markers of replicative senescence (CD28-CD57+) and proliferation (Ki67+) in both the aviremic and viremic group compared with healthy children (Figs. 3B, C), indicating a possible effect of HIV-1 infection on the senescence of these cells. Moreover, the presence of higher viral load in the viremic group produced a slightly higher replicative senescence phenotype when compared with aviremic group (P = 0.052) (Fig. 3B), indicating that viral load favors the senescence of memory CD8 T cells. Additionally, the viral load in the viremic group produces as well an increase in the activated HLADR+CD38+ memory subset compared with aviremic and healthy children. Additionally, the viral load in the viremic group produces as well an increase in the activated HLADR+CD38+ memory subset compared with aviremic and healthy children."
Daz, Laura PhD; Mendez-Lagares, Gema; Correa-Rocha, Rafael PhD; Pacheco, Yolanda Mara PhD; Ferrando-Martnez, Sara PhD; Ruiz-Mateos, Ezequiel PhD; Pozo-Balado, Mara del Mar del; Leon, Juan Antonio MD, PhD; Gurbindo, Mara Dolores MD, PhD; Isabel de Jose, Mara MD, PhD; Leal, Manuel MD, PhD; Munoz-FernAndez, M. Angeles MD, PhD
Laboratorio de InmunoBiologa Molecular, Instituto de Investigacion Sanitaria Gregorio Maranon, Hospital General Universitario Gregorio Maranon, C/Doctor Esquerdo, Madrid, SpainLaboratorio de Inmunovirologa, Instituto de Biomedicina de Sevilla (IBIS), Servicio de Enfermedades Infecciosas, Hospital Universitario Virgen del Roco. Avda. Manuel Siurot s/n, Sevilla, SpainDepartamento de Bioqumica Clnica, IBIS/CSIC/Universidad de Sevilla, Hospital Universitario Virgen del Roco, Sevilla, SpainUnidad de Enfermedades Infecciosas PediAtricas. Hospital Universitario Virgen del Roco, Sevilla, SpainSeccion de Inmuno-Pediatra, Hospital General Universitario Gregorio Maranon. C/Doctor Esquerdo, Madrid, SpainUnidad de Enfermedades Infecciosas PediAtricas. Hospital Universitario La Paz, P° de la Castellana, Madrid, Spain.
Abstract
Background: CD8 T cells are crucial in the immune responses against HIV infection, but HIV-infected adults suffer a naive CD8 T-cell depletion and accelerated senescence caused by chronic antigen stimulation. Although HIV-infected children preserve a better immune reconstitution capacity their CD8 responses are defective. We wanted to know, whether HIV vertical transmission produces a premature aging of the CD8 population due to antigen exposition to HIV from birth and persistent chronic activation.
Methods: We conducted a multicentre cross-sectional study that compared vertically HIV-infected children with detectable (viremic) or undetectable (aviremic) viral load and age-matched healthy children. Using multiparameter flow cytometry, we studied within the CD8 population the frequencies of naive, memory, effector memory (effector memory), and TemRA subsets and measured markers of senescence, activation, and proliferation in these cells.
Results: We found that naive subset in viremic children was markedly decreased and had a replicative senescence phenotype. Furthermore, viremic children showed increased frequencies of memory, TEM and TemRA CD8 T cells, with a more activated and replicative senescence phenotype. We found that HIV-infected children with undetectable viral load have an increased senescence in memory and effector CD8 T cells, but the frequencies and phenotype of the CD8 subsets analyzed are comparable to healthy children.
Conclussions: Our study shows that CD8 T cells of HIV-infected children have a more senescent phenotype when compared with age-matched healthy children. Interestingly enough, our results support the importance of maintaining undetectable viral load in HIV-infected children to avoid the premature ageing and dysfunction of CD8 T cells.
INTRODUCTION
Nowadays mother-to-child transmission of HIV remains a major world-wide health problem, with a yearly estimation of 370,000 new vertically HIV-infected children.1 In resource-rich countries, the use of highly active antiretroviral therapy (HAART) has decreased the HIV-1 vertical transmission to the rate as low as 0.99%2 and reduced infant mortality by around 75%. HIV-infected children survive to adolescence and adulthood, and the challenges of providing HIV care are evolving into the challenges of providing both acute and chronic life-long care. Continuing treatment for life is problematic, given the limitations of the available drugs, the long-term toxicity of antiretroviral therapy, and adherence issues.3 In addition, persistent antigen exposure to HIV have been associated to hyperactivation of immune system, degradation of immune responses, and AIDS progression.4,5
It is well known that persistent exposure to HIV-1 antigens accelerates the aging of the immune system in HIV-infected individuals.6,7 Such aging process, similarly to what occurs in the elderly non-HIV-infected people,8,9 involves several homeostatic alterations including phenotypic changes and loss of functionality of CD8 T cells,10,11 which are crucial for the control of the infection.
Antigenic stimulus in HIV infection promotes differentiation, proliferation and senescence of effector CD8 T cells,10 and also induces the proliferation of highly differentiated antigen-specific CD4 and CD8 memory T cells.7,12 These CD8 responses lead to the exhaustion of CD8 T-cell pool,13 which is followed by a replenishment of the consumed CD8 T cells from the naive CD8 T-cell pool.14 However, HIV-infected adults have reduced levels of naive CD8 T cells15 that are not able to replace old senescent CD8 T cells.6 One of the major changes that occur in HIV-viremic individuals is the accumulation of CD28-CD57+ T cells, particularly within the CD8 population.10 It has been suggested that this replicative senescent phenotype16 or clonal exhaustion17 of HIV-specific CD8 T cells may underlie the inability of T-cell immunity to suppress the virus adequately.10
Children possess a preserved thymic function, and HIV-infected children have a better capacity to reconstitute the immune system than adults.18 However, studies conducted in perinatally HIV-infected children report decreased detection and smaller responses of CD8 T cells than observed in HIV-infected adults.19 Although it has been demonstrated that HIV-1-infected adults undergo the same immunologic changes that healthy elderly persons,20,21 there are very few studies about the immunosenescence in HIV-infected children.22,23 Natural history of HIV infection in vertically HIV-infected children differs markedly from adults.24 It is unknown if these HIV-infected children, who are in contact with the HIV since their birth and who suffer from a persistent chronic activation of their immune system from birth, could suffer from a premature "aging", and how it could affect the immune responses.
Therefore, our aim was to perform an extensive study of the phenotype, and notably immunosenescence features, of different CD8 T-cell subsets and the possible influence of the virological status of vertically HIV-infected children in the degree of senescence.
RESULTS
Characteristics of the Vertically HIV-Infected Children
Fifty-seven HIV-1 vertically infected children and 32 noninfected healthy children were studied. The HIV-infected children were divided into 2 groups as follows: 32 aviremic children with undetectable viral load (<50 copies/mL) and a median age of 14.0 (IQR: 10.3-17.8) and 25 viremic children with viral load >50 copies per milliliter and a median age of 14.0 (IQR: 12.5-16). Children included in the viremic group had viral load higher than 50 copies per milliliter during the last 6 months previous to the sample obtained for this study. Children with undetectable viral load (<50 copies/mL) during the last 6 months previous to the study were included in the aviremic group. The median viral load (IQR) in viremic group was 1730 (260-13600) copies/mL. Median of CD4 T-cell absolute counts and percentages were higher in aviremic than viremic group (Table 1). In contrast, median CD8 T-cell percentage was higher in viremic than aviremic group (Table 1).
Influence of Viral Load and Age in the Frequency of Naive CD8+ T Cells
We studied the percentage of naive (CD45RA+CD27+) CD8 T cells in all children. The results showed that the frequency of naive CD8 T cells was significantly diminished in the viremic group compared with aviremic group (P < 0.001) and non-HIV healthy children (P < 0.001) (Fig. 1A). Naive CD8 T cells of viremic group had an increased frequency of CD28-CD57+ cells in comparison to healthy children (P < 0.001), which were associated to a replicative senescence phenotype. Furthermore, the phenotype of CD8-naive subset in this viremic group is also more senescent (P = 0.007) and proliferative (P = 0.007) than in the aviremic group (Figs. 1B, C). In summary, in aviremic children, the frequency, proliferation, and senescence of naive CD8 T cells are not modified in comparison to healthy children. However, viremic group shows significant alterations in these parameters.
Next, we studied if the percentage of naive CD8 T cells in HIV-infected and non-HIV-infected children was correlated with the age. Our results indicate that frequency of naive CD8 T cells decreases with age in healthy children (P = 0.001, r = 0.579) (Fig. 2A). In contrast, both aviremic and viremic groups did not show significant correlation between age and percentage of naive CD8 T cells (P = 0.534, r = 0.120 and P = 0.698, r = 0.083, respectively) (Figs. 2B, C). In addition, healthy children shown a positive correlation between naive CD8 and naive CD4 T cells (P < 0.001, r = 0.830) (Fig. 2D), which was not found in vertically HIV-infected patients (Figs. 2E, F). Summing up, in healthy children both naive CD4 and CD8 T cells decrease with age, but percentages of naive CD8 T cells in HIV-infected children are altered and the frequency does not depend on the age.
Influence of HIV Infection and Viral Load in Memory CD8+ T Cells
A significant increase of memory CD8 T cells was observed in viremic group compared with healthy children (P < 0.001) (Fig. 3A), but this accumulation of memory CD8 T cells was not observed in aviremic children showing a frequency comparable to healthy group. Memory T-cell subset showed a higher markers of replicative senescence (CD28-CD57+) and proliferation (Ki67+) in both the aviremic and viremic group compared with healthy children (P < 0.001) (Figs. 3B, C), indicating a possible effect of HIV-1 infection on the senescence of these cells. Moreover, the presence of higher viral load in the viremic group produced a slightly higher replicative senescence phenotype when compared with aviremic group (P = 0.052) (Fig. 3B), indicating that viral load favors the senescence of memory CD8 T cells. Additionally, the viral load in the viremic group produces as well an increase in the activated HLADR+CD38+ memory subset compared with aviremic and healthy children.
METHODS
Study Subjects
We carried out a cross-sectional study on a cohort of 57 vertically HIV-infected children with a median age of 14.0 [interquartile range (IQR): 11.5-17] from the Hospital Universitario Virgen del Roco, Hospital General Universitario Gregorio Maranon, and Hospital Universitario La Paz, Spain. At the same time, 32 healthy noninfected children as a control group median age of 16.0 (IQR: 10.3-17) were studied.
Fresh samples from patients were partially processed by the Laboratory from Instituto de Biomedicina de Sevilla (Seville) and the Pediatric HIV BioBank integrated in the Spanish AIDS Research Network (RED RIS).25 Informed consent was obtained from parents or legal tutors for each children, and protocols were approved by institutional ethical committees.
Analysis of Clinical Parameters
Absolute numbers, percentages of CD4 and CD8 T cells were determined in fresh whole blood using the flow cytometer-500 (Beckman Coulter Inc, CA), according to the manufacturer instructions.
Plasma HIV-1 RNA levels was measured by quantitative polymerase chain reaction (PCR) (COBAS Ampliprep/COBAS Taqman HIV-1 test, Roche molecular systems), according to the manufacturer instructions. The detection limit was 50 copies per milliliter.
Isolation of CD8 T Cells
Fresh peripheral blood mononuclear cells (PBMC) were isolated through a Ficoll-Hypaque gradient centrifugation. For CD8 T-cell isolation, PBMC from HIV-infected or healthy children, were labeled with CD8 microbeads by direct magnetic labeling. Positive fraction was isolated on magnetic columns (MiltenyBiotec, Germany). The purity was >99.9%.
Monoclonal Antibodies and Subpopulation Analysis
To determine the frequency of different subpopulations in the same sample, we isolated CD8 T cells from PBMC. The monoclonal antibodies used for the analyses were as follows: Ki67 CD45-RO-PECy7 (Becton Dikinson, San Jose, CA); CD57-FITC, CD45-RA-FITC, CD28-PE, Ki67-PE, CD38-PE, CD45-RA-ECD, HLA-DR-ECD, CD27-PC5 (Beckman Coulter, FL). CD8 T-cell subsets were defined as follows: naive (CD45RA+CD27+), memory (CD45-RO+CD27+), TEM (CD45RO+CD27-) and effector memory RA+ (TemRA) (CD45RA+CD27-) by flow cytometry. The accuracy of these phenotypes has recently been reported.26 To identify activated T cells, HLADR and CD38 markers were used. Senescent CD8 T cells were characterized as CD28-CD57+. Proliferation levels were determined by intracellular immunostaining with Ki67. Cells were permeabilized using the fixation/permeabilization staining set (eBioscience) according to the manufacturer instructions. Apoptosis in CD8 T-cell subsets was studied using Annexin V and 7-ADD staining.
Cells were analyzed using a Beckman-Coulter flow cytometry-500 cytometer with CXP software that provides automatic daily monitoring of system performance thorough Levy-Jennings plots. The analysis was realized with Expo32 software (Coulter Corporation) program. Dot plot cursor settings were based on isotype control staining and samples were stained with single-color antibodies in the presence of isotype control antibodies. For data analysis the following populations were gated in sequence. The first gate was SSC vs FSC to identify the cell population of interest. The rest of the gates were selected based on the expression or the markers CD45RA, CD45RO, and CD27. At least, 50.000 events were collected for each sample.
Statistical Analysis
All continuous variables were expressed as median (IQR), and categorical variables as the number of cases and percentages. Differences among categorical variables were analyzed using the χ2 test. The Mann-Withney U test was used to analyze differences between continuous variables. Correlations between quantitative parameters were explored using the Spearman rho correlation coefficient. Statistical analysis was performed using the Statistical Package for the Social Sciences software (SPSS 15, Chicago, IL). A P value <0.05 was considered statistically significant.
Influence of HIV Infection and Viral Load on TEM and TemRA CD8 T Cells
We analyzed the influence of HIV infection on the frequency of TEM (CD45RO+CD27-) and a terminally differentiated effector subset named TemRA (CD45RA+CD27-) in CD8 T cells. Aviremic and healthy children shown a similar proportion of both TEM and TemRA CD8 T cells and a significant increase of these subsets was observed only in children with uncontrolled viral load (Figs. 4A, D). Both TEM and TemRA cells showed a more senescent phenotype in HIV patients than controls (Figs. 4B, E). Differences were significant in both aviremic group (P = 0.008 and P < 0.001 for TEM and TemRA cells, respectively) and in the viremic group (P = 0.012 and P = 0.006 for TEM and TemRA cells, respectively). We also observed that activation of TEM and TemRA cells, indicated by HLA-DR expression, was increased only in viremic and not in aviremic children when compared with healthy group (Figs. 4C, F). Moreover, in the viremic group, we also found a higher expression of HLADR but also a higher proportion of other activated subsets such as HLADR+CD38+ cells compared with aviremic and healthy children.
Additionally, we analyzed the degree of apoptosis in these CD8 subsets, and we observed that in both viremic and aviremic group have a higher apoptosis in TemRA cells compared with healthy children. (P = 0.006 and P = 0.003, respectively). However, we did not find any differences in the apoptosis degree of effector memory subsets between all studied groups.
Summing up, our results suggest that regardless the viral load, viremic and aviremic children show an increased senescence in TEM and TemRA subsets in comparison to healthy children and a higher degree of apoptosis in these subsets.
DISCUSSION
This is the first study that shows an exhaustive analysis of CD8 T-cell subsets in vertically HIV-infected children. For this purpose, we measured not only the frequency, but also the grade of senescence, proliferation, and activation of these cells. Our findings provide evidences of premature immunosenescence in HIV-infected children, notably in viremic children, and reveal the importance of maintaining the HIV-infected children with undetectable viral load.
We investigated the correlation of CD8 T cells with viral load or age. There are conflicting findings regarding the association between CD8 T-cell responses with viral load or disease progression in infants27; with some studies showing a positive,19 negative,28,29 or absence of correlation.30 These contradictory findings are likely to be due to phenotypical differences in the CD8 T-cell population depending on virological or immunological factors of the patients.
Aging has shown to increase the presence of senescence markers as CD57 in T lymphocytes leading to impairment in the functionality of these cells.31 We hypothesize that permanent antigenic stimulus and immune hyperactivation in HIV-infected children could induce a premature ageing of the immune system similar to the effect of age in healthy subjects. This hypothesis was confirmed by our results showing that HIV-infected children have a higher frequency of senescence markers in the CD8 population than age-matched healthy children. Moreover, our results also demonstrated that presence of uncontrolled viral load in HIV-infected children favors the accumulation of CD8 T cells with a memory and effector phenotype and increases the degree of senescence in these CD8 T cells.
Studies show that HIV-1 preferentially infects actively dividing cells and in children there is an accelerated cell division.22 This may provide an explanation for some of the distinctive pathogenic features of the HIV disease in children, including higher viral loads and more rapid disease progression. Moreover, most therapeutic regimens have failed to achieve a complete suppression of HIV-1 replication in children32,33 and transient viremia was found among children on HAART.34 All HIV-infected children enrolled in this study were under HAART, and presence of viral load in viremic group was mainly due to the absence of good adherence in these children.
Firstly we analyzed the naive CD8 population that is crucial to maintain the pool of CD8 T cells. The loss of naive CD8 T cells has been related to impaired cytotoxic responses during HIV infection.35,36 In healthy donors these naive population decrease with age probably due to the impairment in the thymic production of new T cells.35,37 However, this correlation with age disappears in the vertically HIV-infected group. We have previously described that HIV infection impairs thymic function,38 and thymic production of new T cells in HIV-infected children is negatively correlated with viral load.39,40 This fact would explain that, from the birth and along the infection, children with higher viral load have lower frequencies of naive CD8 T cells than aviremic or healthy children. Percentage of naive CD8 T cells from aviremic group and healthy group were similar, confirming that the level of viral load is the factor responsible for the decrease of naive CD8 T cells and not the fact of being infected by HIV. The absence of correlation between age and naive CD8 T cells in aviremic group reflects the increased turnover of these cells along the infection for the replenishment of consumed CD8 cells.41 Naive CD8 and naive CD4 frequencies were correlated in healthy children, which is probably associated to the thymic production of T cells, but this correlation disappears in HIV-infected children. Exhaustion and replenishment of CD8 pool in HIV-infected subjects are very different from the CD4 pool leading to different dynamics and frequencies in the CD4 or CD8 population. The high turnover of CD8 T cells in HIV-infected children41,42 without an adequate replenishment of new CD8-naive cells will consume quickly the pool of naive CD8 T cells. Moreover, our results indicate that naive CD8 T cells in viremic children are more senescent and show a more replicative phenotype than in aviremic and healthy children. Increased turnover of the CD8 population in presence of antigenic stimulus and absence of an adequate thymic output of naive cells lead to the ageing of naive CD8 pool that it is reflected by an increase in this replicative senescence phenotype. Summing up, HIV-mediated impairment of thymic production could induce a deficiency in the replenishment of the naive CD8 pool in patients with uncontrolled viral load comparable to the age-mediated impairment observed in older healthy children. HAART-mediated control of viral load preserves the pool of naive CD8 T cells in aviremic children and avoids the HIV-mediated premature ageing of these cells.
On the other hand, viremic group had a higher frequency of memory TEM and TemRA CD8 T cells with a more activated phenotype than aviremic or healthy group. TemRA cells represent a very terminally differentiated end-stage effector cells that are unable to proliferate on restimulation and they have a potent antiviral activity,43 and play a vital role to control the viremia at early stages of the infection.44 Moreover, there has been found a relationship between absolute numbers of HIV-1-specific CD8+ T cells expressing CD57 with a TemRA phenotype.44 In addition, a skewed in T-cell differentiation from central memory T cells to TEM have been described in HIV-infected adults and children.45 These facts could explain the increased frequency in these effector subsets of CD8 T cells that agree to previous work in HIV-infected children.23 The presence of an active HIV replication even in aviremic children, due to the residual output of HIV particles from the reservoirs,46,47 could produce the observed increase in the proliferation of memory cells, notably in the viremic group. HIV-infected children show a phenotype of memory TEM and TemRA subpopulations more senescent than the phenotype in healthy group. These results indicate that as described in adults,7 HIV infection induces a premature ageing of the effector CD8 pool. The presence of senescence markers was increased in aviremic children when compared with healthy children, as consequence to the HIV exposition from birth. However, the frequency of all CD8 subsets studied is preserved in this aviremic group and level of senescence was significantly lower than in children with uncontrolled viral load. These data clearly indicate that the control of viral load can reduce dramatically the premature ageing in the CD8 T-cell population. The senescence or premature ageing in these effector populations that are crucial in the HIV-specific immune responses will compromise the viability of these cells leading to the exhaustion and the depletion of the CD8 T-cell pool described in the progression of the disease.48
Summing up, our results clearly show that HIV infection in children produces a premature ageing of the CD8 T-cell population, notably in children with higher values of viral load. The continuous immune activation of these cells is likely responsible for the appearance of senescence CD8 T-cell markers and the increased rates of apoptosis observed in HIV-infected subjects. In addition, uncontrolled viral load in viremic group support the accumulation of memory TEM and TemRA cells with a more senescent and activated phenotype. This could indicate that the immune system of viremic children could be subject to greater pressure to eliminate the virus and therefore have a higher proportion of terminally differentiated cells with a senescent phenotype. Senescent CD8 T cells persist due to their resistance to apoptosis and they occupy progressively the memory T-cell pool restricting the repertoire of the remaining T cells.11 Although CD8 T cells have relative resistance to apoptosis, other authors have found that the CD57+CD8+ T cells were more prone to undergo activation-induced cell death after mitogen stimulation.10 Therefore, the presence of HIV (reservoirs or detectable viral load) could be the responsible stimuli for the activated TemRA subset to enter into apoptosis.
The impairment in thymic production of new T cells associated to the infection is likely responsible of the decreased frequency of naive CD8 T cells, whereas memory cells are rising in response to HIV and are associated with increased TemRA cells.49 For most of the subsets analyzed, aviremic group had a phenotype more similar to healthy children than to the viremic group. This fact suggests that the presence of viral load induces a more replicative senescence phenotype in the CD8 population, and ART-mediated control of viral load could reduce dramatically the premature ageing of this population preserving a CD8 T-cell phenotype comparable with healthy children.
At the light of these results, we emphasize the importance of maintaining undetectable viral load in HIV-infected children to avoid a premature ageing and impairment of the CD8-mediated immune responses. Early identification of vertically HIV-infected patients who are likely to experience therapeutic failure is very important as it could promote the initiation of alternative successful therapy with the objective of preserve the immune responses and controlling the progression of the disease.
| |
| |
| |
|
|
|