| |
Scientists Discover New Gene That Affects Clearance of Hepatitis C Virus........A variant upstream of IFNL3 (IL28B) creating a new interferon gene IFNL4 is associated with impaired clearance of hepatitis C virus
|
| |
| |
Download the PDF here
Download the PDF here
.........Scientists have discovered a gene that interferes with the clearance of hepatitis C virus infection. They also identified an inherited variant within this gene, Interferon Lambda 4 (IFNL4), that predicts how people respond to treatment for hepatitis C infection.
......Compared to rs12979860, ss469415590 is more strongly associated with HCV clearance in individuals of African ancestry, although it provides comparable information in Europeans and Asians.......It has been reported that rs12979860 predicts early viral kinetics in HCV-infected patients receiving IFN-α-free treatment52. This genotype has also been associated with the response to IFN-α-based treatment of chronic hepatitis B virus (HBV) infection in some studies53, 54. Furthermore, IFN-α therapy is used for a number of other clinical conditions, including some forms of cancer55. Thus, therapeutic inhibition of IFNL4 might represent a novel biological strategy for the treatment of HCV and HBV infection and possibly other diseases, and IFNL4 genotype could be used to select patients for this therapy.
...........By analyzing data from hepatitis C-infected African-Americans and European-Americans participating in clinical studies, the authors found that the presence of the IFNL4 protein is associated with poorer clearance and response to treatment than the form that does not produce IFNL4. The deletion variant is more common in people of African ancestry, which helps partially explain why African-Americans have a lower response to current hepatitis C treatments than patients of Asian and European ancestry.
..........We have identified a new inducible human protein-coding gene, IFNL4, which is related to but distinct from known IFNs and other class 2 cytokines. The 179 amino-acid ORF of the IFNL4 transcript is created by a common deletion frameshift allele of ss469415590, which is a dinucleotide variant strongly linked with rs12979860. In individuals of African ancestry, the IFNL4-generating ss469415590[∼G] allele is superior to the rs12979860[T] allele in predicting poorer response to pegIFN-α/RBV treatment of CHC. Within IFNL4, we identified three nonsynonymous variants, rs73555604 (p.Cys17Tyr), rs142981501 (p.Arg60Pro) and rs11764844 (p.Pro70Ser), which are present on haplotypes with the ss469415590[∼G] allele. The effects of these variants on IFNL4 biological function and their impact on HCV clearance in different populations should be further explored.
.......We assessed the associations of ss469415590 and rs12979860 with HCV clearance in 1,436 African-American and 1,480 European-American individuals from 4 studies. In Virahep-C32 and HALT-C33, we evaluated the response to pegIFN-α/RBV therapy in individuals with CHC (Supplementary Table 5). There were differences in the rates of sustained virological response (SVR) among the subjects from these studies, which reflect well-known ancestry-related differences in response to treatment and the differing selection criteria for these clinical trials: Virahep-C (European-American), 52%; Virahep-C (African-American), 28%; HALT-C (European-American), 18%; and African-American, 7%. We evaluated spontaneous HCV clearance in injection drug users enrolled in two studies, UHS34 and ALIVE35 (Supplementary Table 6). The decrease in the amount of HCV RNA during the first 28 d of treatment is a powerful early predictor of ultimate treatment response that is strongly associated with rs12979860 genotype36, 37. In African-American Virahep-C participants, the decline in HCV RNA levels after 28 d of treatment was more strongly associated with ss469415590 genotype than with rs12979860 genotype (P = 0.015, difference in mean values; Fig. 3 and Supplementary Table 7). In the same study, we observed a stronger association for ss469415590 than for rs12979860 with other measures of treatment response (week 24 response, end-of-treatment response and SVR; Table 2), although these differences did not reach statistical significance. The association pattern was similar in African-American individuals from the HALT-C study, with a stronger association for ss469415590 than for rs12979860 (week 20 response, end-of-treatment and SVR; Table 2). Spontaneous HCV clearance in African-Americans was evaluated using the area under the receiver operating curve (AUC). In UHS participants, the AUC value was greater for ss469415590 (0.62) than for rs12979860 (0.58) (Table 3). In the ALIVE study, the AUC values were similar for rs12979860 (0.64) and ss469415590 (0.64) (Table 3).
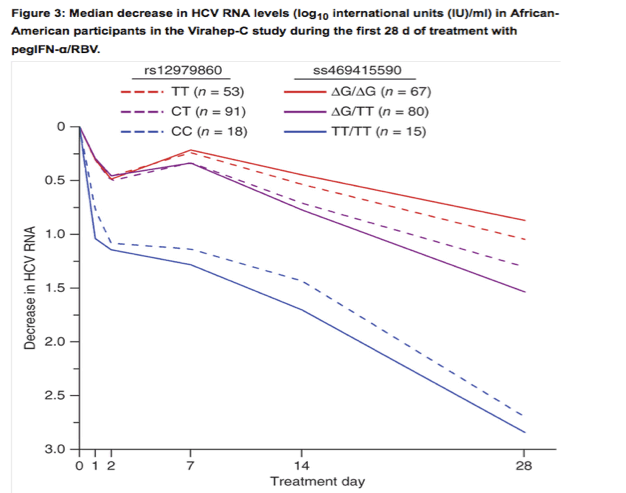
P = 0.015 for the mean differences in HCV RNA levels at day 28 for each of the three genotype groups at ss469415590 relative to the respective rs12979860 genotype groups.
--------------------------------------------
Scientists Discover New Gene That Affects Clearance of Hepatitis C Virus
Jan. 9, 2013 - Scientists have discovered a gene that interferes with the clearance of hepatitis C virus infection. They also identified an inherited variant within this gene, Interferon Lambda 4 (IFNL4), that predicts how people respond to treatment for hepatitis C infection.
The results of this study, by investigators at the National Cancer Institute (NCI), part of the NIH, and their collaborators at NIH and other institutions, were published online in Nature Genetics on Jan. 6, 2013.
Chronic infection with hepatitis C virus is a cause of liver cirrhosis and liver cancer. Up to 80 percent of people who are acutely infected with hepatitis C fail to clear the virus and develop chronic hepatitis C infection, and of these, approximately 5 percent develop liver cancer. Individuals of African ancestry do not respond as well to current treatments of hepatitis C infection compared to patients of European or Asian ancestry.
Previously, results from genome-wide association studies (GWAS) identified common inherited genetic markers that were associated with response to hepatitis C virus treatment and spontaneous clearance of the infection. Those markers are located on chromosome 19 near a known interferon gene, IFNL3 (IL28B). However, molecular investigations into IFNL3 did not explain the GWAS association with spontaneous virus clearance or treatment response. To find the new gene, the investigators used a technology involving RNA sequencing on human liver cells treated to mimic hepatitis C virus infection.
"By using RNA sequencing we looked outside the box to search for something beyond what was already known in this region. We hit the jackpot with the discovery of a new gene. It is possible that other important genes may be discovered using this approach," said co-lead investigator Ludmila Prokunina-Olsson, Ph.D., of the Laboratory of Translational Genomics in NCI's Division of Cancer Epidemiology and Genetics (DCEG).
The researchers found that the IFNL4 region harbors a variant that is found in two alternative forms. One form, called deltaG, results in a deletion in one of the four bases that comprise DNA. The change creates an alteration known as a frameshift, which produces the IFNL4 protein, while the form without the deletion does not produce IFNL4. By analyzing data from hepatitis C-infected African-Americans and European-Americans participating in clinical studies, the authors found that the presence of the IFNL4 protein is associated with poorer clearance and response to treatment than the form that does not produce IFNL4. The deletion variant is more common in people of African ancestry, which helps partially explain why African-Americans have a lower response to current hepatitis C treatments than patients of Asian and European ancestry.
"Our work fulfills several promises of the genomic era," said NCI's Thomas R. O'Brien, M.D., Infections and Immunoepidemiology Branch, DCEG. "One, a better understanding of biology; two, personalized medicine; and three, new potential treatments. We deliver immediately on the first two. We've identified a new gene that may help us better understand response to viral infection and the new genetic marker may transition to clinical practice because it predicts treatment outcome for African-American patients better than the current genetic test. For the third, the INFL4 protein may be used as a novel therapeutic target for hepatitis C virus infection, and possibly other diseases."
The new gene belongs to what is now a family of four interferon-lambda protein-encoding genes, three of which were discovered more than ten years ago (IFNL1, IFNL2 and IFNL3) The mechanism by which the IFNL 4 protein impairs hepatitis C virus clearance remains unknown. Further studies will explore molecular function of this novel protein in normal and disease conditions.
This study was conducted collaboratively with the National Institute of Diabetes and Digestive and Kidney Diseases at NIH, as well as the U.S. Food and Drug Administration, and a number of universities and research institutions. Funding was provided by NCI grant Z01 CP005782.
-----------------------------------------
A variant upstream of IFNL3 (IL28B) creating a new interferon gene IFNL4 is associated with impaired clearance of hepatitis C virus
Nature Genetics Jan 6 2013
Ludmila Prokunina-Olsson1, Brian Muchmore1, Wei Tang1, Ruth M Pfeiffer2, Heiyoung Park3, Harold Dickensheets4, Dianna Hergott1,5, Patricia Porter-Gill1, Adam Mumy1, Indu Kohaar1, Sabrina Chen6, Nathan Brand1, McAnthony Tarway1, Luyang Liu1, Faruk Sheikh4, Jacquie Astemborski7, Herbert L Bonkovsky8, Brian R Edlin9,10, Charles D Howell11, Timothy R Morgan12,13, David L Thomas7,14, Barbara Rehermann3, Raymond P Donnelly4 & Thomas R O'Brien5
ABSTRACT
Chronic infection with hepatitis C virus (HCV) is a common cause of liver cirrhosis and cancer. We performed RNA sequencing in primary human hepatocytes activated with synthetic double-stranded RNA to mimic HCV infection. Upstream of IFNL3 (IL28B) on chromosome 19q13.13, we discovered a new transiently induced region that harbors a dinucleotide variant ss469415590 (TT or ∼G), which is in high linkage disequilibrium with rs12979860, a genetic marker strongly associated with HCV clearance. ss469415590[∼G] is a frameshift variant that creates a novel gene, designated IFNL4, encoding the interferon-λ4 protein (IFNL4), which is moderately similar to IFNL3. Compared to rs12979860, ss469415590 is more strongly associated with HCV clearance in individuals of African ancestry, although it provides comparable information in Europeans and Asians. Transient overexpression of IFNL4 in a hepatoma cell line induced STAT1 and STAT2 phosphorylation and the expression of interferon-stimulated genes. Our findings provide new insights into the genetic regulation of HCV clearance and its clinical management.
Introduction
More than 3% of the world population is infected with HCV1. Up to 80% of acutely infected individuals fail to clear the virus and develop chronic hepatitis C (CHC)2, with as many as 5% eventually progressing to liver cancer3. Success of CHC treatment with pegylated interferon-α with ribavirin (pegIFN-α/RBV) depends on HCV genotype and reaches 50-80% in patients of European ancestry but only 30% in patients of African ancestry. Adding a direct-acting antiviral (DAA) agent to this regimen increases the success rate but may result in adverse effects from pegIFN-α/RBV4 and DAA treatment. If this treatment fails, there is an increased risk of the selection of resistant HCV strains that may compromise future treatment options5, 6.
Recent genome-wide association studies (GWAS) have identified the rs12979860 and rs8099917 SNPs on chromosome 19q13.13 near the IFNL3 gene (formerly known as IL28B) as variants associated with both spontaneous HCV clearance7, 8 and response to pegIFN-α/RBV treatment7, 8, 9, 10, 11. Within this region reside the three interferon-λ genes, IFNL1, IFNL2 and IFNL3 (formerly IL29, IL28A and IL28B, respectively), which encode the type III IFNs12, 13. Type I IFNs (IFN-α and IFN-ß) and type III IFNs (IFN-λs) induce antiviral activity and suppress HCV replication in vitro14, 15 and in vivo16 through activation of the JAK-STAT pathway and upregulation of interferon-stimulated genes (ISGs)14, 17, 18. The molecular phenotype of this genetic association remains unclear. The GWAS markers have not consistently been associated with hepatic IFNL3 mRNA expression19, 20, 21, and a nonsynonymous IFNL3 variant, rs8103142 (encoding a p.Arg70Lys alteration), which is in strong linkage disequilibrium (LD) with rs12979860 in all HapMap populations (r2 = 0.8-1.0), does not seem to affect the function of the IFNL3 protein22.
Discussion
We have identified a new inducible human protein-coding gene, IFNL4, which is related to but distinct from known IFNs and other class 2 cytokines. The 179 amino-acid ORF of the IFNL4 transcript is created by a common deletion frameshift allele of ss469415590, which is a dinucleotide variant strongly linked with rs12979860. In individuals of African ancestry, the IFNL4-generating ss469415590[∼G] allele is superior to the rs12979860[T] allele in predicting poorer response to pegIFN-α/RBV treatment of CHC. Within IFNL4, we identified three nonsynonymous variants, rs73555604 (p.Cys17Tyr), rs142981501 (p.Arg60Pro) and rs11764844 (p.Pro70Ser), which are present on haplotypes with the ss469415590[∼G] allele. The effects of these variants on IFNL4 biological function and their impact on HCV clearance in different populations should be further explored.
Analysis of the genomic sequences of 45 species for which the sequence of the IFNL4 region is available in the UCSC Genome Browser showed that the unfavorable IFNL4-generating ss469415590[∼G] allele is an ancestral variant present in all species. The existence of IFNL4 protein could be predicted only in the genomes of macaques (marmosets and rhesus), orangutans, chimpanzees and humans (Supplementary Fig. 9). The beneficial insertion ss469415590[TT] allele seems to be a recently derived variant, which became common in all human populations (93% frequency in Asians, 68% frequency in Europeans and 23% frequency in Africans, according to HapMap samples), suggesting positive selection for this allele. The introduction of frameshifts is considered to be an evolutionary mechanism for the rapid emergence of new proteins49, 50, but, in this case, an insertion allele that abrogates IFNL4 seems to have been selected during evolution.
We found that the IFNL4 protein of 179 amino acids induces STAT1 and STAT2 phosphorylation, activates the ISRE-Luc reporter and ISGs, and generates antiviral response in hepatoma cells. The mechanisms by which IFNL4 induces these responses but nevertheless impairs HCV clearance are currently under investigation. IFNL4 and IFNL3 have similar residues in the area that is known to interact with the primary receptor of IFNL3 (IFNLR1) but differ in the region of IFNL3 that interacts with the second chain of the IFNL receptor complex, IL10R2. Thus, it is possible that IFNL4 activates JAK-STAT signaling through a unique receptor complex consisting of IFNLR1 and a currently undefined second receptor chain or that IFNL4 functions as a decoy cytokine competing with type III IFNs for the binding of IFNLR1. We also found that the IFNL4-caused preactivation of interferon signaling prevents further activation by type I and type III IFNs. We used an allele-specific mRNA expression assay (Supplementary Fig. 10) and explored endogenous IFNL4 expression in PHHs, where it was induced by polyI:C, IFN-α (but not IFNL3) treatment and in vitro infection with HCV (Supplementary Fig. 11). However, no IFNL4 mRNA expression was induced by polyI:C, IFN-α or IFNL3 in several transformed cell lines that carry the ss469415590[∼G] allele (HepG2, HeLa, 293T, A549 and HH29). Experiments aimed at elucidating the triggers of IFNL4 expression in diverse conditions and cell types and its receptor components are ongoing and may provide greater insight regarding its mechanism of action.
Previous studies found that individuals with CHC who carry rs12979860[T], which marks the ss469415590[∼G] allele, before treatment have somewhat higher hepatic expression of ISGs but poorer ISG response to pegIFN-α/RBV treatment19, 41, 42, 43, 44. The rs12979860[T] variant has also been associated with lower HCV RNA levels in the absence of treatment9, 51. Our in vitro experiments in HepG2 cells showed that IFNL4 induced activation of ISGs, which was not further enhanced by exogenous treatment with IFN-α or IFNL3. Taken together, these data suggest that IFNL4 induces weak expression of ISGs that provides an antiviral response that might lower the HCV load, although it also reduces the responsiveness to type I and type III IFNs that is needed for efficient HCV clearance.
It has been reported that rs12979860 predicts early viral kinetics in HCV-infected patients receiving IFN-α-free treatment52. This genotype has also been associated with the response to IFN-α-based treatment of chronic hepatitis B virus (HBV) infection in some studies53, 54. Furthermore, IFN-α therapy is used for a number of other clinical conditions, including some forms of cancer55. Thus, therapeutic inhibition of IFNL4 might represent a novel biological strategy for the treatment of HCV and HBV infection and possibly other diseases, and IFNL4 genotype could be used to select patients for this therapy.
Results
A genetic variant creates IFNL4, a novel interferon protein
We performed sequencing of mRNA (RNA-seq) in primary human hepatocytes (PHHs) treated with polyinosinic:polycytidylic acid (polyI:C), which is a synthetic mimic of double-stranded HCV RNA. The PHH sample was from a liver donor who was heterozygous for rs12979860 (C/T) and uninfected with HCV. Hepatocytes were treated with polyI:C for 0, 1, 2, 4, 8 or 24 h, and induction of the IFN-λ genes (IFNL1, IFNL2 and IFNL3) was confirmed by TaqMan expression analysis before performing RNA-seq. An analysis of RNA-seq data that focused on a 150-kb region centered on rs12979860 showed concordance with the TaqMan expression results-there was no expression of IFN-λ genes without polyI:C treatment, and these genes were induced after 2-24 h of activation by polyI:C (Fig. 1a).
We also observed transient activation of a novel transcribed region upstream of IFNL3, with the highest levels of expression detected at 2 and 4 h after treatment (Fig. 1b). Analysis of paired-end RNA-seq reads identified one major splice junction site. Using this common sequence as a starting point for 5' rapid amplification of cDNA ends (5'RACE), we mapped a transcription start site, followed by a unique protein translation start site 277 bp downstream. Within exon 1 (at amino acid 22), we detected a novel compound dinucleotide variant, denoted ss469415590 (TT>∼G), comprised of a one-base insertion or deletion (indel) polymorphism (loss of T, rs67272382) and a one-base substitution variant (T>G, rs74597329). Using polyI:C-stimulated PHHs from five additional liver donors and the primers described in Supplementary Table 1, we cloned and annotated ten individual transcripts created by a combination of the ss469415590 alleles and inclusion of several alternative exons (Fig. 1c,d; NCBI accession numbers are presented in Supplementary Table 2). The location of these novel transcripts 3 kb upstream of and in the same orientation as IFNL3 raised the possibility that they were alternatively spliced forms of IFNL3 or fusions. However, the presence of a CTCF transcriptional insulator site23, 24 between the two transcribed regions (Fig. 1b), the results of the RACE experiments and the inability to generate an RT-PCR product covering IFNL3 and the novel transcribed region confirmed the independence of these loci. Despite high overall similarity with a genomic region upstream of IFNL2, the novel transcripts and ss469415590 are specific for the region upstream of IFNL3 (Supplementary Fig. 1).
Of the ten novel transcripts, four were interrupted by premature stop codons and, thus, are likely to be eliminated by nonsense-mediated mRNA decay25. The remaining six transcripts were predicted to produce full-length proteins of 143 amino acids (p143) and 124 amino acids (p124) from transcripts with the ss469415590[TT] allele and 179 amino acids (p179), 170 amino acids (p170), 131 amino acids (p131) and 107 amino acids (p107) from transcripts with the ss469415590[∼G] allele (Fig. 1c). A global protein BLAST search found homology only for p179, with 29.1% amino-acid identity and 40.8% amino-acid similarity with IFNL3. However, the p179 and IFNL3 cDNA sequences were not similar enough to be aligned using the BLAST bl2seq tool. On the basis of its protein sequence homology with type III IFNs (Table 1), the International Society of Interferon and Cytokine Research (ISICR) and the nomenclature committee of the Human Genome Organization (HUGO) designated p179 as interferon λ4 protein (IFN-λ4, IFNL4). IFNL3 and p179 (IFNL4) proteins are most related within the sequences that correspond to the A and F helices of IFNL3, which constitute the core area for the interaction of IFNL3 and other type III IFNs with their primary receptor, IFNLR1 (also known as IL28R1). However, IFNL4 differs in the area corresponding to the D helix of IFNL3, which is the area of interaction of type III IFNs with the second chain of the IFN-λ receptor complex, IL10R2 (Fig. 2)26, 27, 28, 29.
Association of genetic variants in IFNL4 with HCV clearance
The GWAS markers rs12979860 and rs8099917 are located 367 bp downstream (intron 1) and 4 kb upstream of ss469415590, respectively. Analysis of data from the HapMap Project30 (Supplementary Table 3) and the 1000 Genomes Project31 (Supplementary Table 4) showed that the IFNL4-creating ss469415590[∼G] allele is perfectly correlated with the unfavorable rs12979860[T] allele in Asians (Han Chinese in Beijing, China (CHB) and Japanese in Tokyo, Japan (JPT) sets, r2 = 1.00) and well correlated in Europeans (Utah residents of Northern and Western European ancestry (CEU) set: HapMap, r2 = 0.92; 1000 Genomes Project, r2 = 0.83). In Africans, however, this correlation is only moderate (Yoruba from Ibadan, Nigeria (YRI) set: HapMap, r2 = 0.71; 1000 Genomes Project, r2 = 0.65), even though rs12979860 is the best surrogate for ss469415590 of all markers within the 100-kb region. Correlation between ss469415590 and rs8099917 was high in Asians (r2 = 0.91) and moderate in Europeans (r2 = 0.44) but very low in Africans (r2 = 0.008) (Supplementary Table 3).
We assessed the associations of ss469415590 and rs12979860 with HCV clearance in 1,436 African-American and 1,480 European-American individuals from 4 studies. In Virahep-C32 and HALT-C33, we evaluated the response to pegIFN-α/RBV therapy in individuals with CHC (Supplementary Table 5). There were differences in the rates of sustained virological response (SVR) among the subjects from these studies, which reflect well-known ancestry-related differences in response to treatment and the differing selection criteria for these clinical trials: Virahep-C (European-American), 52%; Virahep-C (African-American), 28%; HALT-C (European-American), 18%; and African-American, 7%. We evaluated spontaneous HCV clearance in injection drug users enrolled in two studies, UHS34 and ALIVE35 (Supplementary Table 6). The decrease in the amount of HCV RNA during the first 28 d of treatment is a powerful early predictor of ultimate treatment response that is strongly associated with rs12979860 genotype36, 37. In African-American Virahep-C participants, the decline in HCV RNA levels after 28 d of treatment was more strongly associated with ss469415590 genotype than with rs12979860 genotype (P = 0.015, difference in mean values; Fig. 3 and Supplementary Table 7). In the same study, we observed a stronger association for ss469415590 than for rs12979860 with other measures of treatment response (week 24 response, end-of-treatment response and SVR; Table 2), although these differences did not reach statistical significance. The association pattern was similar in African-American individuals from the HALT-C study, with a stronger association for ss469415590 than for rs12979860 (week 20 response, end-of-treatment and SVR; Table 2). Spontaneous HCV clearance in African-Americans was evaluated using the area under the receiver operating curve (AUC). In UHS participants, the AUC value was greater for ss469415590 (0.62) than for rs12979860 (0.58) (Table 3). In the ALIVE study, the AUC values were similar for rs12979860 (0.64) and ss469415590 (0.64) (Table 3).
Virahep-C, HALT-C and UHS also enrolled European-American participants. In these subjects, ss469415590 and rs12979860 showed similar associations for both treatment-induced and spontaneous HCV clearance (Supplementary Tables 7-10). Taken as a whole, our results show that, in African-American individuals, ss469415590 is a better marker than rs12979860 for predicting response to pegIFN-α/RBV treatment of CHC and possibly for spontaneous HCV clearance, whereas these variants are similarly informative in European-Americans.
By sequencing IFNL4 in 270 HapMap samples, we annotated 3 nonsynonymous variants, rs73555604 (p.Cys17Tyr) in exon 1 and rs142981501 (p.Arg60Pro) and rs117648444 (p.Pro70Ser) in exon 2, as well as 4 synonymous variants, rs150891559 (p.Ala11Ala) and rs4803221 (p.Ser30Ser) in exon 1 and rs12971396 (p.Ser149Ser) and rs137902769 (p.Ser175Ser) in exon 5 (Fig. 2, Supplementary Fig. 2 and Supplementary Tables 11 and 12). On the basis of a haplotype analysis of 16 markers from the 8-kb IFNL3-IFNL4 region, we identified 8 markers that captured all haplotypes present in the HapMap sets (Supplementary Table 13). These eight markers were also tested in European-American and African-American individuals from Virahep-C. In all populations, the unique favorable haplotype included the ss469415590[TT] allele, which eliminates the IFNL4 protein. The unfavorable ss469415590[∼G] allele was found on a number of haplotypes, including two haplotypes that were reported as being neutral in Europeans despite carrying the unfavorable rs12979860[T] allele38, 39; these two haplotypes included minor alleles of either of the rs73555604 and rs11764844 nonsynonymous variants. It is possible, therefore, that these variants modify the risk in carriers of the unfavorable ss469415590[∼G] allele and are the source of the haplotype heterogeneity previously reported in Europeans38, 39; however, data from Virahep-C are too sparse to confirm this theory (Supplementary Table 13).
IFNL4 induces expression of ISGs
We evaluated the functional properties of the six novel protein isoforms created by alleles of ss469415590. For an analysis of 45 signaling pathways, HepG2 hepatoma cells were transiently transfected with expression constructs for all six isoforms or treated with recombinant IFN-α, IFNL3 or IFNL4 (with the latter two produced in the sfs9 baculoviral expression system). Only transfection with construct expressing IFNL4, as well as treatment with IFN-α or IFNL3, induced activation of an interferon-stimulated response element luciferase reporter (ISRE-Luc), which contains STAT1- and STAT2-binding sites responsive to type I and type III IFN signaling and the IRF1 reporter (Fig. 4a). These results were validated in HepG2 cells transiently (Fig. 4b), as well as stably (Fig. 4c), expressing ISRE-Luc reporter constructs. The effect was comparable when the cells were transfected with constructs expressing IFNL4 that generated proteins with either a Halo or a Flag tag (Supplementary Fig. 3). Similarly, only transient transfection with the construct expressing IFNL4 decreased HCV RNA replication in hepatoma cells stably expressing a subgenomic luciferase-expressing hepatitis C virus replicon40 (Fig. 4d) and induced STAT1 and STAT2 phosphorylation (Fig. 4e). Transfection with IFNL4 activated the ISRE reporter in HepG2 and 293T cells but not in HeLa cells (Supplementary Fig. 4).
Recombinant IFNL4 protein expression was detectable in the cells and cell lysates of transfected HepG2 and 293T cells by confocal imaging and protein blots, respectively, with antibodies specific for IFNL4 and tag proteins (Supplementary Figs. 5 and 6). We also detected weak expression of recombinant IFNL4 in the medium of transfected HepG2 cells but not 293T cells (Supplementary Fig. 7). In polyI:C-stimulated PHHs from liver donors not infected with HCV, the endogenous expression of IFNL4 protein was detected by confocal imaging in carriers of the unfavorable ss469415590[∼G] allele but not in a homozygous carrier of the favorable ss469415590[TT] allele (Fig. 5). In hepatocytes from the donor heterozygous for ss469415590[∼G], we detected endogenous expression of IFNL4 in cells treated with polyI:C and after in vitro infection with the JFH1 HCV strain (Supplementary Fig. 6d). In fact, in hepatocytes from one of these donors (∼G/TT genotype), we observed low IFNL4 expression, even without polyI:C treatment or in vitro HCV infection (at the time point at 0 h). Although preliminary, these results suggest that IFNL4 might be expressed in conditions unrelated to HCV infection.
To further explore the functional consequences of IFNL4 expression, we performed RNA-seq in HepG2 cells transiently transfected with empty vector or construct expressing IFNL4 (Supplementary Fig. 8a) and found that the top canonical pathways induced by IFNL4 were related to the activation of interferon signaling (Supplementary Fig. 8b). We validated the RNA-seq results by quantitative RT-PCR (qRT-PCR) analysis and showed that IFNL4 induced the expression of many ISGs in a pattern similar to that induced by IFN-α and IFNL3 (Supplementary Fig. 8c). Previously, many of these ISGs have been shown to be expressed at higher levels in liver biopsies taken before treatment from HCV-infected patients who do not respond to pegIFN-α/RBV treatment; these individuals tend to carry the unfavorable genotypes at rs12979860 and rs8099917 (ref. 19,41,42,43,44), which mark the ss469415590[∼G] allele that produces IFNL4. To mimic this clinical phenotype, we transfected HepG2 cells with empty vector or IFNL4 expression constructs and/or treated cells with 10 ng/ml of recombinant IFN-α or IFNL3. In these samples, we validated the RNA-seq data by qRT-PCR analysis and showed that IFNL4 induced expression of selected ISGs (STAT1, ISG15, IFIHI1-MDA5, OAS1, MX1 and DHX58-RIG-I) in a pattern similar to that induced by IFN-α and IFNL3 (Supplementary Fig. 8d). Furthermore, treatment with IFN-α or IFNL3 of cells already expressing IFNL4 did not induce additional activation of ISGs (Supplementary Fig. 8d). Some genes known as markers of HCV-induced liver damage, such as the chemokine CCL5 (also known as RANTES)45 and the proto-oncogene FOS46, 47, 48, were induced by IFNL4 but not by IFNs (Supplementary Fig. 8d), suggesting a functional role for IFNL4 distinct from the roles of IFN-α and IFNL3.
|
|
| |
| |
|
|
|