| |
Gut - Brain Connection - Signals from the gut microbiota to distant organs in physiology and disease
|
| |
| |
Download the PDF here
Download the PDF here
Nature Medicine Oct 6 2016
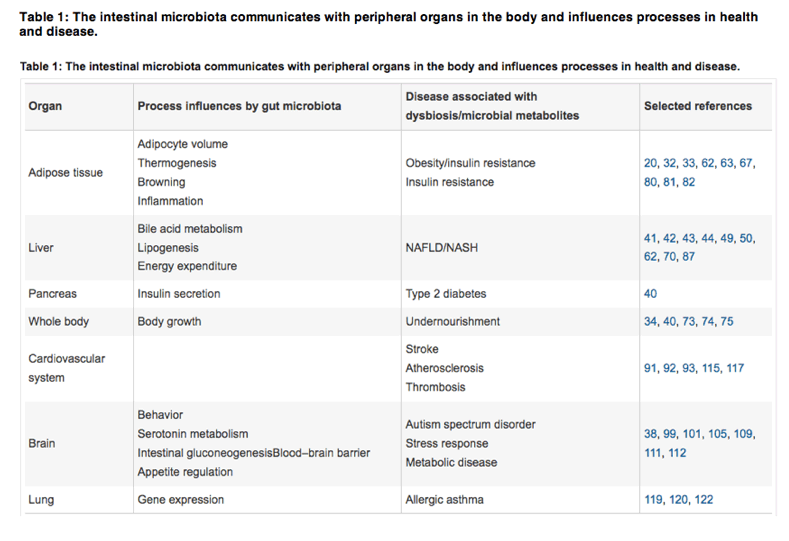
There are also accumulating data suggesting that many human diseases have their origin in distorted gut microbiota composition-or potentially, in microbial metabolites that signal to distant organs (Table 1). Here we discuss recent findings of how the gut microbiota signals to peripheral organs distant from the gut, and how this communication affects physiology and disease.
"Germ-free or antibiotic-treated mice exhibit immature microglia with impaired immune response to bacterial and viral products"
Introduction
There are more microbial cells in the gut as human cells in the body1. Approximately 1,200 different bacterial species have been identified in at least the same amount of the human gut microbiota, and each individual is host to a distinct set of at least 160 species in the gut2, 3, 4, 5, 6. The collective microbial genome encodes 500 times more genes than the human genome3, 6, 7, and so it is tempting to consider human genes as noise in the storm of microbial signals. Recent data suggest that microbial signals modulate crucial functions of the healthy human body, ranging from host metabolism to brain function. There are also accumulating data suggesting that many human diseases have their origin in distorted gut microbiota composition-or potentially, in microbial metabolites that signal to distant organs (Table 1). Here we discuss recent findings of how the gut microbiota signals to peripheral organs distant from the gut, and how this communication affects physiology and disease.
The gut microbiota
In humans, microbial density increases from the proximal to the distal end of the intestine and comprises a biomass of 1.5-2.0 kg, dominated by strictly anaerobic Bacteria3. Archaea, Eukarya and viruses are also present, but their relevance for human health has been less studied.
Although the vast majority of the gut microbial community is composed of only five phyla (Bacteroidetes, Firmicutes, Actinobacteria, Proteobacteria and Verrucomicrobia), there is considerable diversity on the species level and their relative abundances. Key members of Firmicutes include the genera Clostridium, Lactobacillus and Ruminococcus, as well as the butyrate producers Eubacterium, Fecalibacterium and Roseburia. Members of Bacteroidetes are known to be efficient degraders of dietary fiber and include the genera Bacteroides, Prevotella and Xylanibacter. Bifidobacterium is a major genus within Actinobacteria, and several taxa are used as probiotics. Proteobacteria includes Escherichia and Desulfovibrio, whereas Verrucomicrobia so far includes only the mucus-degrading genus Akkermansia.
The composition of the gut microbiota is influenced by genetic and environmental factors starting early in life (Box 1, also recently reviewed elsewhere8). It has been postulated that each individual can be grouped into one of three bacterial clusters called enterotypes, which are defined by the relative abundance of the genera Bacteroides, Prevotella or Ruminococcus9. Although the presence of enterotypes has been debated10 and is probably not as discrete as originally suggested, it is clear that members of the gut microbiota co-occur and rely on each other's metabolic activities11. By contrast, competition for similar environmental conditions and nutrients restricts bacterial colonization and leads to niche competition within or between bacterial species11, 12, 13.
Gut microbiota analyses in humans are often based on fecal material, which is easily accessible. Yet microbiota composition varies along the intestinal tract and differs even between the intestinal lumen and mucosa-adherent bacteria that reside in the mucus4. Because of oxygen diffusion from the epithelium, even aerobic bacteria have been detected in mouse crypts14. Thus, analysis of fecal microbiota composition is a valuable tool and could serve as a biomarker, but it might not accurately reflect the microbes that are in closest contact with the host. Moreover, as we will discuss in this Review, many of the physiological effects attributed to the gut microbiota are caused by their metabolites. Because different microbes might produce the same metabolites and those small molecules are less restricted in their spatial diffusion15, the presence or absence of a single bacterial species is valuable but not sufficient for understanding the detailed interaction between the microbiota and the host.
EXCERPTS
Undernourishment.
Evidence is accumulating that the gut microbiota is involved in many human diseases (Fig. 3). Altered microbial communities as a result of both undernourishment and obesity have been suggested to contribute to the pathogenesis of these diseases, given that the metabolic phenotype of these disorders can be transferred to germ-free mice, provided the relevant diet, upon fecal transplant67, 68, 69, 70, 71.
As the first dietary encounter, breast milk is a source rich in human milk oligosaccharides (HMOs), which resist degradation in the upper small intestine and nourish the colonic gut bacteria as soluble fiber72. Total and sialylated HMOs were reduced in mothers with infants whose growth was severely stunted73, and supplementation of sialylated bovine milk oligosaccharides promoted growth in gnotobiotic mice and piglets colonized with microbiota from infants with stunted growth73. No growth effect was observed in germ-free mice, demonstrating a microbiota-dependent effect. Given these findings, it will be notable to investigate whether sialylated HMOs promote the expansion of distinct microbiota that are beneficial for the host, or whether microbial degradation products such as sialylated monosaccharides or sialic acid are the beneficial metabolite promoting host growth.
Furthermore, undernourished children were found to harbor immature gut microbiota74, and the presence of a mature community or the addition of Ruminococcus gnavus and Clostridium symbiosum were sufficient to rescue growth impairment in mice harboring the immature microbiota from undernourished children70. Although the underlying molecular signals are unknown, a recent study highlighted that germ-free mice have stunted growth on a diet that was sufficient to promote adequate growth in colonized mice75. The stunting was associated with reduced activity of the somatotropic growth hormone (GH)- insulin-like growth factor-1 (IGF1) axis, a major driver of postnatal growth, as revealed by reduced IGF1 serum levels and decreased expression of IGF1 in the liver in germ-free as compared to convetionally raised mice75. Mono-colonization of undernourished juvenile GF mice with a selected Lactobacillus plantarum strain recovered somatotropic axis signaling and systemic growth. However, mechanisms underlying the increase in IGF signaling remain to be identified, as does the relevance of this pathway in humans whose growth is stunted. But supplementing nutrition therapy with specific probiotic commensals might enhance dietary interventions used to treat undernutrition in the future.
Atherosclerosis.
Exploiting the diet-microbiota pathway has recently been suggested for the treatment of atherosclerosis. An unhealthy alliance between host and gut microbiota converts choline, found in seafood, cheese and eggs, as well as carnitine, found in red meat, into TMAO, a toxic compound associated with atherosclerosis and cardiovascular disease91, 92, 93 (Fig. 1). The primary step is the formation of trimethylamine (TMA) from choline by means of microbial TMA lyases. Consequently, the inhibition of TMA lyases is a promising step toward reducing risk for atherosclerosis. Indeed, 3,3-dimethyl-1-butanol (DMB), a structural analog of choline, was found to inhibit microbial TMA production in vitro and in mouse experiments94. Furthermore, oral DMB application reduced macrophage-foam-cell formation and aortic-root atherosclerotic plaque development in atherosclerosis-prone Apoe-/- mice. Thus, drugging the bugs95 to treat diseases of the host is a promising concept that needs to be validated in human diseases, especially when complex dietary interactions might complicate the situation. Remarkably, the authors detected the identified inhibitor DHB in various foods and drinks, including balsamic vinegars, cold-pressed extra virgin olive oils and red wines. Whether such a combination reaches a sufficiently high DHB concentration in the human gut and so reduces the risk of atherosclerosis needs to be investigated.
Gut microbial signals in the gut-brain axis
Brain morphology.
Investigating the influence of gut microbiota on brain morphology is a challenging task in humans, and thus data are generated mostly in germ-free mice. These have alterations in the structural integrity of the amygdala and hippocampus when compared to colonized mice96. Moreover, mice lacking microbiota display increased hippocampal neurogenesis97 and hypermyelination of the prefrontal cortex98. Although mechanistic data are lacking so far, it is evident that the gut microbiota has an effect on the structure of the (mouse) brain. Colonization of germ-free mice with defined bacterial groups or isolated species could reveal whether distinct microbes or a complex community are required to alter the morphology of brain regions. Likewise, it would be intriguing to determine whether the same bacteria are responsible for morphological alterations in different regions of the brain. Yet, given that access to the brain is tightly controlled by the blood-brain barrier (BBB), it is likely that a small and specific set of bacterial metabolites modulates brain morphology.
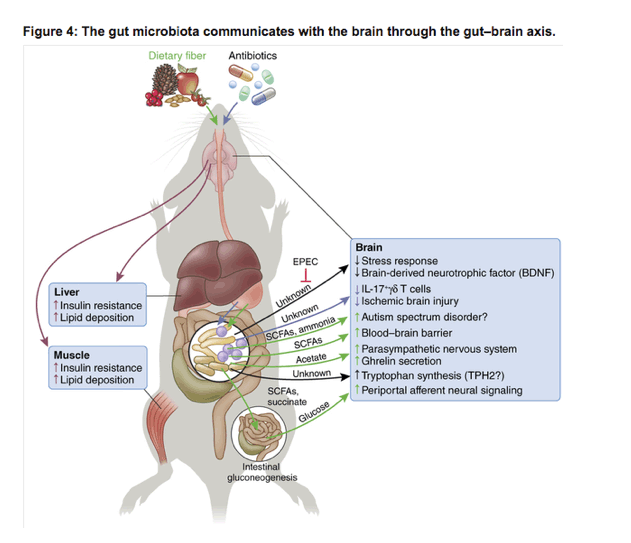
The gut microbiota is also crucially involved in modulating the BBB. Germ-free mice have a more permeable BBB than do conventionally raised (CONV-R) mice, a phenotype that was reversed after colonization with SCFA-producing Clostridium tyrobutyricum or B. thetaiotaomicron99 (Fig. 4). However, future research is required to identify the molecular mechanism of how butyrate or other SCFAs exhibit a BBB-modulating effect, especially whether epigenetic or GPCR signaling is involved and whether the results can be translated to the human body.
Young germ-free mice (GF, mice that lack microbiota) have increased stress response and increased expression of brain-derived neurotrophic factor (BDNF) in comparison to colonized mice. Colonizing GF mice with enteropathogenic Escherichia coli (EPEC) aggravated the stress response101 by an unknown mechanism (black arrows). Antibiotic treatment alters the gut microbiota composition (violet arrows), which reduces the recruitment of IL-17+γδ T cells to the meninges, thus reducing ischemic brain injury115. Dietary fiber (green arrows) is fermented by the colonic gut microbiota into short-chain fatty acids and succinate. Increased levels of the SCFAs acetic, propionic, butyric, isobutyric, valeric and isovaleric acid, as well as ammonia were observed in children with autism spectrum disorder109. A causative role of the microbiota, however, remains yet to be determined. SCFAs tighten the BBB99, while the SCFA acetate activates the parasympathetic nervous system, leading to increased secretion of the hormone ghreline, leading to insulin resistance and lipid deposition in liver and muscle (red arrows)40. SCFAs butyrate and propionate as well as succinate activate intestinal gluconeogenesis, which leads to central metabolic improvement via gut-brain neural circuits38. The gut microbiota also modulates tryptophan synthesis in the brain105, probably through a tryptophan hydroxylase 2 (TPH2)-dependent manner.
Although access to the brain is controlled by the BBB, microglia are responsible for immune defense in the brain. Germ-free or antibiotic-treated mice exhibit immature microglia with impaired immune response to bacterial and viral products100. Moreover, microglia morphology and distribution were different between colonized and microbiota-deficient mice. Notably, supplementing germ-free mice with a SCFA mix could restore most alterations in microglial function. Similarly, Ffar2-/- knockout mice, which lack the SCFA receptor GPR43, displayed alterations in microglia morphology but not in cell densities. Thus, microbiota-derived SCFAs are probably responsible for most, yet not all, effects of gut microbial modulation of microglia function.
Given that Ffar2 was not expressed in the central nervous system (CNS)100, GPR43 activation needs to occur in the periphery, but the exact organ or cell type needs to be identified. In addition, direct inhibition of histone deacetylase by butyrate might also occur in the CNS.
---------------------------
The candidate compound in plasma with an m/z of 76 associated with CVD risks was isolated and unambiguously identified as TMAO using multinuclear nuclear magnetic resonance (NMR), multi-stage mass spectrometry (MSn), liquid chromatography with tandem mass spectrometry (LC/MS/MS) and gas chromatography with tandem mass spectrometry (GC/MS/MS) after multiple derivatization strategies (see Methods, Supplementary Figs 2a-d, and Supplementary Table 2). TMAO, an oxidation product of trimethylamine (TMA), is a relatively common metabolite of choline in animals12, 13. Foods rich in the lipid phosphatidylcholine (PC, also called lecithin), which predominantly include eggs, milk, liver, red meat, poultry, shell fish and fish, are believed to be the major dietary sources for choline, and hence TMAO production14.
Intestinal microflora have a role in TMAO formation from dietary free choline13. We therefore proposed that commensal organisms (gut flora) might also have an obligate role in TMAO formation from dietary PC. To test this, deuterated PC was synthesized whereby the choline-methyl groups were deuterium labelled (that is, d9-PC) and used as isotope tracer for feeding studies. When mice were fed through oral gavage with d9-PC, the time-dependent appearance of the anticipated d9 isotopomer of TMAO was observed in plasma (Fig. 2c).
Interestingly, pre-treatment of mice with a three-week course ofbroad-spectrum antibiotics to suppress intestinal flora completely suppressed the appearance of d9-TMAO in plasma after oral d9-PC administration (Fig. 2c). A similar pattern was observed after oral administration of d9-choline to mice, with d9-TMAO produced in untreated mice, but not in the same mice after a 3-week course of broad-spectrum antibiotics (Supplementary Fig. 7b), or in germ-free mice born sterilely by Caesarean section (Supplementary Fig. 7c). In a final series of studies, mice with suppressed intestinal microflora after antibiotics were placed in conventional cages with normal (non-germ-free) mice to permit intestinal colonization with microbes. After four weeks, repeat oral d9-PC challenge of the now 'conventionalized' mice resulted in readily detectable plasma levels of d9-TMAO (Fig. 2c). Similar results were observed after conventionalization of germ-free mice and oral d9-choline (Supplementary Fig. 7c). Collectively, these results show an obligate role for intestinal microbiota in the generation of TMAO from the dietary lipid PC. They also reveal the following metabolic pathway for dietary PC producing TMAO: PC choline TMA TMAO.
Pdf attached at top of report above
----------------------------
|
|
| |
| |
|
|
|