| |
Brief Report: Differential Effects of Tenofovir, Abacavir, Emtricitabine, and Darunavir on Telomerase Activity In Vitro
|
| |
| |
Download the PDF here
AIDS Jan 1 2016 Stella-Ascariz, Natalia MS; Montejano, Rocio MD; Pintado-Berninches, Laura MS; Monge, Susana MD; Bernardino, Jose I. MD; Perez-Valero, Ignacio MD; Montes, Maria L. MD; Mingorance, Jesus PhD; Perona, Rosario PhD; Arribas, Jose R. MD
There is growing concern about the issue of aging of HIV-infected patients. It is well established that HIV-infected patients have an increased risk for several "non-AIDS" complications (cardiovascular disease, malignancy, liver disease, kidney disease, bone disease, and neurocognitive decline)that are classically associated with the normal aging process......It is also unknown if this accentuated or accelerated aging [from Jules: its accelerated in general, not accentuated depending on the specific comorbidity] is caused by theproinflammatory stateassociated with even well-controlled HIV infection, traditional risk factors (such as smoking) that are more prevalent among HIV-infected people, or other still unknown causes.......Another potential cause of accelerated or accentuated aging in HIV-infected patientscould be telomere shortening caused by antiretroviral drugs.5There is a close association between shortened telomere length (TL) in peripheral blood mononuclear cells (PBMCs) and diseases of aging, including increased cardiovascular diseases and dementia.6,7
------------------------
Neurotoxicity Screening of Antiretroviral
Drugs With Human iPSC-Derived Neurons.....
http://www.natap.org/2016/CROI/croi_148.htm
------------------------
"In our study, we explored if N(t)RTIs could play a role in the aging process of HIV-infected patients by means of inhibition of telomerase activity and whether DRV could have a similar effect. We have found that TFV and ABC, but not FTC, produced a significant dose-dependent decrease of telomerase activity in PHA-activated PBMCs within the therapeutic concentration range in vivo. After 72 hours of treatment, telomerase inhibition caused by TFV was more than double the inhibition caused by ABC: 29% and 12%, respectively. The observed decrease in telomerase activity caused by TFV and ABC was not associated with a decrease in hTERT protein level, or a change in the expression ofhTERTgene or the other genes that code for the subunits of the telomerase/shelterin complexes. Furthermore, we have shown that DRV did not affect telomerase activity,hTERTgene expression, or hTERT protein levels."
TFV and ABC Inhibit Telomerase Activity In Vitro
We analyzed the effect of 3 N(t)RTIs (TFV, ABC and FTC) and 1 PI (DRV) on telomerase activity in PHA-activated PBMCs from healthy volunteers after 72 hours of treatment. We performed 5 independent experiments with all N(t)RTIs and 6 independent experiments with DRV. Telomerase activity was expressed relative to untreated PBMCs.
Of the N(t)RTIs tested, only TFV and ABC induced a significant dose-dependent decrease of telomerase activity within the therapeutic concentration range. Median inhibition induced by TFV at 0.5 and 1 μM was 29% (IQR: 29%-34%; range: 12%-39%;P= 0.042) and 28% (IQR: 28%-41%; range: 25%-47%;P= 0.042), respectively. For ABC at 3 and 10 μM, median inhibition was 12% (IQR: 9%-13%; range: 8%-17%;P= 0.043) and 14% (IQR: 10%-29%; range: 7%-40%;P= 0.043), respectively. Exposure to FTC or DRV did not affect telomerase activity even at concentrations above the therapeutic plasma level range (Fig. 1A).
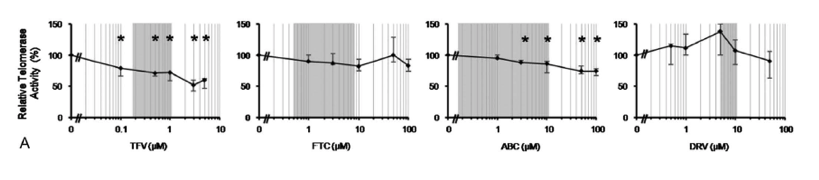
The main objective of our study was to confirm if ABC, FTC and especially TFV, at therapeutic concentrations, inhibit telomerase activity in vitro in activated-PBMCs. Secondly, we assayed if DRV is able to increase telomerase activity in vitro. Finally, we wanted to evaluate the possible impact of these antiretrovirals on mechanisms of telomerase inhibition different from chain termination such as expression of telomerase genes.
There is growing concern about the issue of aging of HIV-infected patients. It is well established that HIV-infected patients have an increased risk for several "non-AIDS" complications (cardiovascular disease, malignancy, liver disease, kidney disease, bone disease, and neurocognitive decline) that are classically associated with the normal aging process......It is also unknown if this accentuated or accelerated aging [from Jules: its accelerated in general, not accentuated depending on the specific comorbidity] is caused by the proinflammatory state associated with even well-controlled HIV infection, traditional risk factors (such as smoking) that are more prevalent among HIV-infected people, or other still unknown causes.......Another potential cause of accelerated or accentuated aging in HIV-infected patients could be telomere shortening caused by antiretroviral drugs.5There is a close association between shortened telomere length (TL) in peripheral blood mononuclear cells (PBMCs) and diseases of aging, including increased cardiovascular diseases and dementia.6,7
The in vivo relevance of the differential effects of TFV, ABC, FTC, and DRV on telomerase activity remains to be elucidated. Although Leeansyah and colleagues - http://jid.oxfordjournals.org/content/207/7/1157.full?sid=800da733-6731-4695-9ce8-15953aa6d1f2 [NRTIs and, specifically tenofovir at therapeutic concentrations, inhibit telomerase activity leading to accelerated shortening of TL in activated PBMCs. The relationship between NRTI, reduced telomerase activity, and accelerated aging requires further investigation in HIV-infected individuals on cART.] - reported that in vivo telomere length was significantly inversely associated with the total duration of treatment with any N(t)RTI, other studies have not found this association. In a substudy of the MONET clinical trial18-
http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0109718 - comparing darunavir/ritonavir monotherapy versus darunavir/ritonavir and 2 N(t)RTIs for maintenance of virological suppression, there were not significant differences between 2 arms after 3 years of follow-up in telomerase activity or mean change per year of telomere length. [Patients in the sub-study had a mean age of 44 years and had received NRTI for a mean of 6.4 years (range 1-20 years). As expected, older patients have significantly shorter TL (p = 0.006), while women had significantly longer TL (p = 0.026). There was no significant association between TL and either the duration of prior NRTI treatment (p = 0.894) or the use of a PI versus NNRTI (p = 0.107). There was no significant difference between patients who continued or ceased NRTI in the mean change/year of TL or telomerase (p = 0.580 and 0.280 respectively).....Factors associated with baseline TL and telomerase activity were analysed by multivariable analysis (Table 2). As expected, older patients has significantly shorter TL (p = 0.006), while women had significantly longer TL (p = 0.026). There was no significant association between TL and either the duration of prior NRTI treatment (p = 0.894) or the use of a PI versus NNRTI (p = 0.107). In the multivariable analysis, TL was significantly longer in PBMC from patients enrolled in the DRV/r monotherapy arm, compared to the DRV/r + 2NRTIs arm (p = 0.008). The reason for this baseline difference was unclear - the longer TL in the DRV/r monotherapy arm was also observed when only men were included in the analysis (p = 0.02). None of the baseline predictors showed significant associations with telomerase activity......In the DRV/r monotherapy arm, the mean (standard deviation, SD) TL (T/S ratio) declined from 1.54 (0.58) at baseline to 1.46 (0.55) at the end of treatment (mean change/year = −0.04). In the DRV/r +2NRTIs arm, the mean TL declined from 1.27 (0.38) at baseline to 1.17 (0.43) at the end of treatment (mean change/year = −0.04). There was no significant difference in the mean change/year of TL between arms (p = 0.580).....In the multivariable analysis, TL was significantly longer in PBMC from patients enrolled in the DRV/r monotherapy arm, compared to the DRV/r + 2NRTIs arm (p = 0.008). The reason for this baseline difference was unclear - the longer TL in the DRV/r monotherapy arm was also observed when only men were included in the analysis (p = 0.02). None of the baseline predictors showed significant associations with telomerase activity.......We then analysed annual changes in TL in PBMC from patients enrolled in the DRV/r +2NRTI arm according to the NRTIs taken. The mean (SD) annual change in TL in patients taking TDF was −0.05 (0.097)/year (n = 23); ABC was −0.05 (0.154)/year (n = 22); and ZDV was −0.03 (0.091)/year (n = 14). There were two patients taking other NRTIs, who were not included in this analysis.]
---------------------
There is growing concern about the issue of aging of HIV-infected patients. It is well established that HIV-infected patients have an increased risk for several "non-AIDS" complications (cardiovascular disease, malignancy, liver disease, kidney disease, bone disease, and neurocognitive decline) that are classically associated with the normal aging process......It is also unknown if this accentuated or accelerated aging is caused by the proinflammatory state associated with even well-controlled HIV infection, traditional risk factors (such as smoking) that are more prevalent among HIV-infected people, or other still unknown causes.......Another potential cause of accelerated or accentuated aging in HIV-infected patients could be telomere shortening caused by antiretroviral drugs.5 There is a close association between shortened telomere length (TL) in peripheral blood mononuclear cells (PBMCs) and diseases of aging, including increased cardiovascular diseases and dementia.6,7
Telomerase is a ribonucleoprotein enzyme complex with a RNA template (TERC), a human telomerase reverse transcriptase (hTERT) subunit, and other regulatory proteins that together with the shelterin complex maintains telomere structure.8 Telomerase adds repetitive TTAGGG sequences to the ends of chromosomes, compensating for the progressive telomeric loss occurring at each cell division. Because of structural and mechanistic similarity to HIV reverse transcriptase, nucleoside/nucleotide reverse transcriptase inhibitors [N(t)RTI] can inhibit telomerase.9 Zidovudine (AZT), stavudine (d4T), didanosine (ddI), and abacavir (ABC) can inhibit telomerase activity in replicating cell lines in vitro, leading to accelerated shortening of TL.10,11 This inhibition is not observed with nonnucleoside reverse transcriptase inhibitors.12 Recently, Leeansyah and collaborators reported that tenofovir (TFV)13 at therapeutic concentrations is a potent inhibitor of telomerase activity, causing telomere shortening in vitro. In Leeansyah et al study, lamivudine (3 TC) and emtricitabine (FTC) also inhibited telomerase activity although only at high concentrations. This finding is surprising because, being cytidine analogues, 3 TC and FTC should not act as chain terminators and therefore other mechanism of telomerase inhibition by 3 TC and FTC could be operating.....Two studies have reported that saquinavir, a protease inhibitor (PI), is able to increase telomerase activity in PBMCs14,15and demonstrated that telomerase up-regulation appeared to be the result of enhanced expression of hTERT in human T leukemia cells in vitro.
TFV and ABC Inhibit Telomerase Activity In Vitro
We analyzed the effect of 3 N(t)RTIs (TFV, ABC and FTC) and 1 PI (DRV) on telomerase activity in PHA-activated PBMCs from healthy volunteers after 72 hours of treatment. We performed 5 independent experiments with all N(t)RTIs and 6 independent experiments with DRV. Telomerase activity was expressed relative to untreated PBMCs.
Of the N(t)RTIs tested, only TFV and ABC induced a significant dose-dependent decrease of telomerase activity within the therapeutic concentration range. Median inhibition induced by TFV at 0.5 and 1 μM was 29% (IQR: 29%-34%; range: 12%-39%; P = 0.042) and 28% (IQR: 28%-41%; range: 25%-47%; P = 0.042), respectively. For ABC at 3 and 10 μM, median inhibition was 12% (IQR: 9%-13%; range: 8%-17%; P = 0.043) and 14% (IQR: 10%-29%; range: 7%-40%; P = 0.043), respectively. Exposure to FTC or DRV did not affect telomerase activity even at concentrations above the therapeutic plasma level range (Fig. 1A).
DISCUSSION
In our study, we explored if N(t)RTIs could play a role in the aging process of HIV-infected patients by means of inhibition of telomerase activity and whether DRV could have a similar effect. We have found that TFV and ABC, but not FTC, produced a significant dose-dependent decrease of telomerase activity in PHA-activated PBMCs within the therapeutic concentration range in vivo. After 72 hours of treatment, telomerase inhibition caused by TFV was more than double the inhibition caused by ABC: 29% and 12%, respectively. The observed decrease in telomerase activity caused by TFV and ABC was not associated with a decrease in hTERT protein level, or a change in the expression of hTERT gene or the other genes that code for the subunits of the telomerase/shelterin complexes. Furthermore, we have shown that DRV did not affect telomerase activity, hTERT gene expression, or hTERT protein levels.
The active forms of TFV, ABC, and FTC compete with the intracellular dATP, dGTP and dCTP pools, respectively, and the incorporation of these nucleotide analogs causes viral DNA chain termination. Telomeres are made up of hexamer repeat TTAGGG sequences. Telomerase binds to the 3' end of the telomeres through its own RNA template and adds TTAGGG polynucleotides to the extreme, whereas the complementary strand is filled in by DNA polymerase later on. Consequently, inhibition of the reverse transcriptase activity of hTERT by the chain termination mechanism would be possible with TFV and ABC but not with FTC. Leeansyah and collaborators13 have previously shown that TFV was the only N(t)RTI that at therapeutic concentrations in vitro significantly inhibited telomerase activity and enhanced shortening of TL. However, in their study, ABC and FTC were also able to inhibit telomerase activity at concentrations above the therapeutic range, but only ABC enhanced shortening of TL. Probably, our study underestimates the inhibition of activity telomerase with ABC and FTC because we performed measurements of telomerase activity after 72 hours of treatment with a single dose of each drug, in contrast to Leeansyah study in which N(t)RTIs were replenished every 48 hours. This difference could account for the lack of effect of FTC on telomerase activity observed in our study. Moreover, another limitation of our study is that we cannot determine whether the main effects of NRTIs were on CD4+ or CD8+ T cells, or specific subsets of T cells or non-T-cell population.
The in vivo relevance of the differential effects of TFV, ABC, FTC, and DRV on telomerase activity remains to be elucidated. Although Leeansyah and colleagues reported that in vivo telomere length was significantly inversely associated with the total duration of treatment with any N(t)RTI, other studies have not found this association. In a substudy of the MONET clinical trial18 comparing darunavir/ritonavir monotherapy versus darunavir/ritonavir and 2 N(t)RTIs for maintenance of virological suppression, there were not significant differences between 2 arms after 3 years of follow-up in telomerase activity or mean change per year of telomere length.
Our results provide more evidence about the inhibition caused by some NRTI on telomerase activity. We have confirmed that TFV and ABC inhibit telomerase activity in activated PBMCs in vitro at therapeutic concentrations and that TFV is the most potent inhibitor. Telomerase activity inhibition caused by N(t)RTIs is probably due to inhibition of hTERT activity leading to chain termination and does not involve changes in expression levels of telomerase genes or hTERT protein. In addition, we did not find that DRV affects telomerase activity, hTERT gene expression, or hTERT protein levels. To our knowledge, ours is the first study showing a lack of implication of hTERT proteins levels and mRNA expression. The in vivo relevance of these findings remains to be elucidated.
|
|
| |
| |
|
|
|