| |
Resistance to Direct Antiviral Agents in Patients With Hepatitis C Virus Infection
|
| |
| |
Download the PDF here
Gastroenterology Feb 2010
Christoph Sarrazin, Stefan Zeuzem
J. W. Goethe-University Hospital, Medizinische Klinik 1, Frankfurt am Main, Germany
Received 12 October 2009; accepted 12 November 2009. published online 14 December 2009.
from Jules: drug resistance as it relates to the new oral HCV drugs, and 20-30 are listed in the table below, is crucial to understand. We do not yet know if resistance will persist after it develops, and it does develop, and lead to cross-resistance to other drugs in the class or prevent response to other drugs in the same class later on, but at this time it is important to consider that it might persist and if it does this needs to be considered in treatment decision making. I one poster at AASLD 2 years ago resistance to telaprevir persisted in several individual patients after up to 3 years, and this may occur with other protease inhibitors or other oral HCV drugs, but an important question is if resistance persists will it prevent response to treatment again with the same drug or class of drug. This has not yet been tested.
"It is important to increase our understanding of mechanisms of resistance mutations, viral breakthrough, and relapse after therapy with DAA agents. Improving our knowledge of these processes will require more precise and earlier characterization of resistance profiles of patients given monotherapy, long-term follow-up data, and studies with combination therapies. Studies also are underway to test the effects of combinations of DAA agents without interferon alfa or ribavirinčthese trials will provide valuable information about the ability of DAA agents to suppress HCV replication without development of resistance or even to eradicate the virus."
"It is not known whether resistant variants can persist in the long term as major or minor quasispecies in nonresponsive patients or those with viral breakthrough or relapse after DAA therapy. Thus, genotypic sequence analysis should be performed in the first days during therapy, directly after viral breakthrough, at the end of treatment, and during short-term and long-term follow-up evaluation. Initial characterization of the HCV resistance profile to DAA agents, using the highly sensitive clonal sequencing method, is mandatory. However, other methods for sequence analysis are available (ie, direct sequencing, TaqMAMA, hybridization assays, restriction enzyme assays, deep sequencing methods). It remains to be established which approach will be most useful to identify DAA agent resistance mutations in clinical practice.65......After termination of treatment with telaprevir the number of wild-type isolates rapidly increased within the first 7-10 days of follow-up evaluation. After 3-7 months no high-level resistance was observed anymore and the majority of resistant variants were replaced by wild-type virus.23 However, in single patients V36 and R155 variants still were detectable at low levels and in a recent report on long-term follow-up evaluation even after up to 3 years in single patients low- to medium-levels of V36 and R155 variants were observed.77......differences between linear tetrapeptide and macrocyclic inhibitors binding to the active site of the NS3 protease have been revealed (Figure 3). However, R155 is the main overlapping position for resistance, and different mutations at this amino acid site within the NS3 protease confer resistance to all protease inhibitors that are currently in advanced clinical development......Several nucleoside inhibitors targeting the active site of the HCV-RNA-dependent RNA-polymerase as well as a number of nonnucleoside inhibitors binding to different allosteric sites of the NS5B polymerase are currently in clinical development......R7128, another nucleoside polymerase inhibitor with antiviral activity in genotype 1-infected patients in phase 1 monotherapy studies,91 currently is being investigated in phase 2 clinical trials in genotype 1-, 2-, and 3-infected patients in combination with pegylated interferon and ribavirin as well as in combination with the protease inhibitor R7227/ITMN191. Also from these studies no viral breakthrough with selection of resistant variants was reported. In vitro, different resistant variants for R7128/NM283 (S282T) and R1626 (S96T) have been selected and cross-resistance was excluded......At least 4 different allosteric binding sites have been identified for inhibition of the NS5B polymerase by nonnucleoside inhibitors (Table 1). A number of mutations conferring resistance to the different chemical classes of nonnucleoside inhibitors have been observed mainly in in vitro studies (Figure 4)......VCH-759, VCH-916, and VCH-222 are 3 other NNI-site 2 inhibitors with antiviral activities in monotherapy studies (Figure 1C).16, 100, 101 For VCH-759 as well as VCH-916 viral breakthroughs with selection of resistant variants conferring high (M423T/V/I) and medium (L419V/M, I482L/V/T, V494A/I) levels of resistance were observed (Figure 4).16, 102 In the majority of patients who experienced a viral breakthrough resistance mutations were found to remain present up to 13 days despite a high HCV replication rate and a reduced replication fitness of the variants in vitro.16.....ABT-333, another palm site inhibitor, showed antiviral activity in patients with genotype 1 infection (Figure 1C) and from in vitro replicon studies C316Y and Y448C were observed as main resistance mutations.106, 107 In addition, S556G was reported to be associated with viral breakthrough in a clinical study (Figure 4).106.....In a single ascending dose study it was shown that inhibition of the NS5A protein with BMS-790052 leads to a sharp initial decline of HCV-RNA concentrations (Figure 1D).108 BMS-790052 is the first NS5A inhibitor binding to domain I of the NS5A protein, which was shown to be important for regulation of HCV replication.109 No clinical data on resistance to this class of drugs have been presented yet and results of multiple dose studies have to be awaited."
Chronic hepatitis C virus (HCV) infection is one of the major causes of cirrhosis, hepatocellular carcinoma, and liver failure that leads to transplantation. The current standard treatment, a combination of pegylated interferon alfa and ribavirin, eradicates the virus in only about 50% of patients. Directly acting antiviral (DAA) agents, which inhibit HCV replication, are in phase 1, 2, and 3 trials; these include reagents that target the nonstructural (NS)3 protease, the NS5A protein, the RNA-dependent RNA-polymerase NS5B, as well as compounds that directly inhibit HCV replication through interaction with host cell proteins. Because of the high genetic heterogeneity of HCV and its rapid replication, monotherapy with DAA agents poses a high risk for selection of resistant variants. We review the parameters that determine resistance, genotypic and phenotypic resistance profiles of DAA agents, and strategies to avoid the selection of resistant variants.
Drug resistance results from variations in drug targets (bacteria, fungi, viruses, tumor cells, and so forth) that affect signal transduction pathways, receptors, drug binding sites, or even host proteins to make the therapeutic agent ineffective. Chronic hepatitis C virus (HCV) infection, which affects more than 180 million patients worldwide and is one of the leading causes of cirrhosis and liver failure, is treated with a combination of pegylated interferon alfa and ribavirin. HCV strains with specific mutations that confer resistance to these drugs have not yet been identified in patients, yet only 50%-60% of patients treated with pegylated interferon alfa and ribavirin achieve a sustained virologic response.1 This low rate of response can be accounted for by several viral and host-related factors.2, 3, 4, 5 Furthermore, because of significant side effects associated with the administration of interferon alfa and ribavirin, combination drug doses and treatment duration are restricted. More effective and better-tolerated treatment options for HCV infection are needed. Directly acting antiviral (DAA) agents that are also named specific, targeted antiviral therapies (STAT-C) for HCV infection are in phase 1-3 trials. We review resistance to DAA agents, focusing on compounds in development such as reagents that target the HCV nonstructural (NS)3 protease, the NS5A protein, and the RNA-dependent RNA polymerase NS5B. We also discuss indirect inhibitors or compounds that inhibit HCV replication by not yet completely resolved mechanisms, such as cyclophilin inhibitors, nitazoxanide, and silibinin (Table 1, Figure 1).
Table 1. Direct Antiviral Drugs in Development for HCV Infection
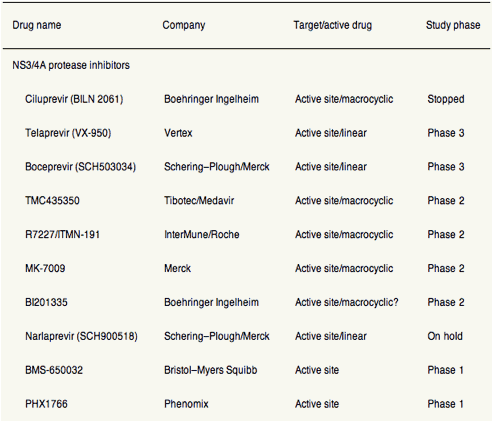
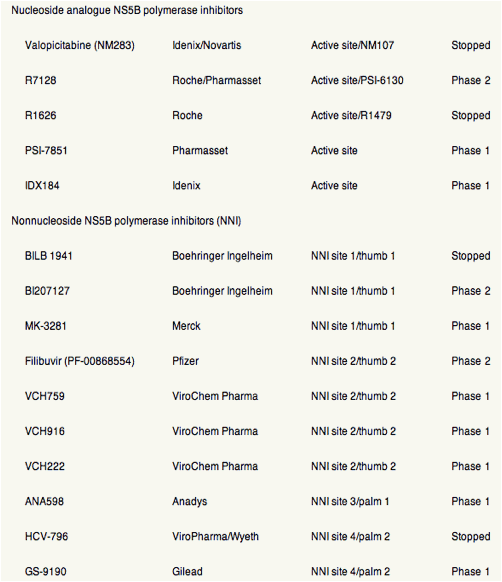
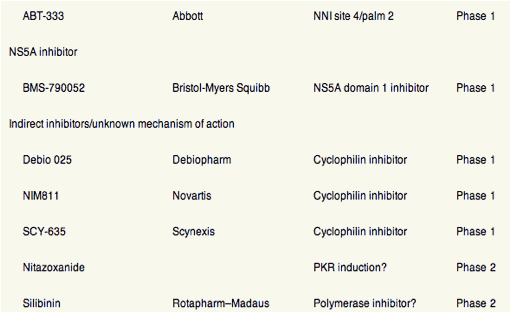
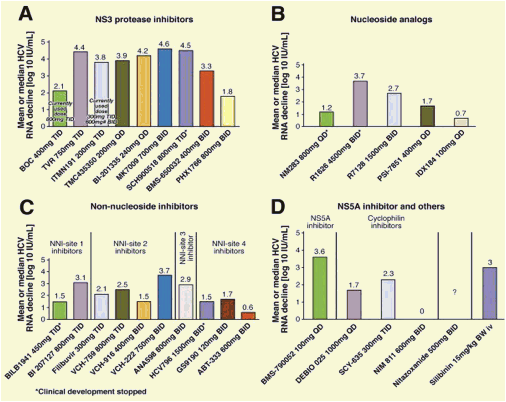
Figure 1. Antiviral activities (mean or median maximum decrease in HCV RNA log10 IU/mL) of DAA agents, as determined from phase 1 monotherapy studies (3-14 days) in patients infected with HCV genotype 1. Different reagents or durations of treatment were not compared directly in these studies. In patients who received the DAA for 7-14 days, the decrease in viral load might not reflect only the activity of the DAA agent but also the emergence of resistant variants. *Clinical development stopped. (A) NS3 protease inhibitors. (B) Nucleoside inhibitors of NS5B. (C) Nonnucleoside inhibitors of NS5B. (D) NS5A inhibitor, indirect inhibitors and agents with unknown mechanism of action. QD, every day, BID, twice a day, TID, three times a day, BW, body weight, #, 900 mg BID stopped from further development due to increase of liver enzymes in single cases.
Parameters That Affect Resistance
Heterogeneity of HCV
HCV has a high rate of turnover; its half-life was estimated to be only 2-5 hours, with the production and clearance of 1010 to 1012 virions per day in an infected patient.6, 7 The HCV-RNA-dependent RNA-polymerase NS5B has poor fidelity and lacks proof-reading activity. Because of this feature and the high replication activity of HCV, a large number of viral variants are produced continuously during infection (error rate, approximately 10-3 to 10-5 mutations per nucleotide per genomic replication).8 The majority of variants produced result in loss of viral replication via loss of function of encoded proteins or lack of tolerance by the host's immune system.9, 10 However, a large number of variants survive; the sum of these different isolates in an infected patient is called viral quasispecies. Typically a dominant strain (wild-type) is detectable within the viral quasispecies along with strains that are present at lower frequencies. The frequencies of HCV isolates depend on their replication efficacies and other known and unknown viral and host factors. During the phylogenetic evolution of HCV, at least 6 different genotypes and more than 100 subtypes have developed that are characterized by large differences in nucleotide sequence (31%-33% and 20%-25%, respectively).11 Thus, the development of HCV therapies is challenged by the different genotypes and subtypes of virus in different patients and also the variation in viral isolates in each patient. Given the heterogeneities in viral sequence, not all drugs will be equally effective for all HCV genotypes, subtypes, and isolates. However, variations are not distributed equally throughout the HCV genome; compounds that target more conserved regions (ie, the active site of the RNA-dependent RNA polymerase) have a better chance of efficacy in patients infected with different genotypes and subtypes.
HCV genotypes and resistance
Many DAA compounds that are being tested in clinical studies are designed to inhibit HCV genotype 1 isolates and the majority of studies have been performed in patients infected with HCV genotype 1. However, telaprevir, an inhibitor of the HCV NS3 protease that targets the active site of the enzyme, has been given subsequently also to patients infected with genotypes 2, 3, and 4. In these patients, viral load decreased by 3.9, 0.5, and 0.9 log10 IU/mL, respectively, after 2 weeks of monotherapy, compared with 4.4 log10 IU/mL in genotype 1-infected patients.12, 13, 14 Also, boceprevir has been reported to have weaker antiviral activity in patients infected with genotypes 2 or 3 and similar results are to be expected for other HCV protease inhibitors. Different levels of activity against different HCV genotypes also are expected from nonnucleoside NS5B inhibitors based on results from in vitro studies; little clinical data are available.15 Administration of VCH-759, for example, which binds the thumb 2 site to inhibit the polymerase NS5B, for 10 days led to a mean maximal decrease in HCV RNA log10 IU/mL of 2.5 in genotype 1-infected patients, whereas no viral decline was observed in a genotype 6-infected patient.16
The active site of NS5B is highly conserved between different genotypes, so nucleoside inhibitors could have similar levels of activity in patients infected with different genotypes. However, there are no data on the efficacy of monotherapy with nucleoside analogs against different genotypes. An interim analysis of data from a placebo-controlled trial of patients infected with HCV genotypes 2 or 3 who did not respond to interferon therapy reported increased rapid virologic response rates after 4 weeks of triple therapy with pegylated interferon alfa, ribavirin, and the nucleoside analogue R7128, compared with standard combination therapy (without the NS5B inhibitor). So, it appears that NS5B inhibitors have antiviral activity in patients with HCV genotypes 2 or 3.17
No clinical data are available on the efficacy of NS5A inhibitors in patients with HCV non-genotype 1, but given the relatively high level of heterogeneity in the NS5A protein, differences in antiviral activities against different genotypes are expected.
Agents that act on host proteins, such as cyclophilin inhibitors, would be predicted to have similar levels of antiviral activity against all HCV genotypes. However, monotherapy with the cyclophilin inhibitor Debio-025 was significantly less effective in patients infected with genotypes 1 or 4 compared with genotypes 2 or 3; viral loads decreased by 2.5 and 4.4 log10 IU/mL, respectively, after 4 weeks monotherapy with 1000 mg Debio-025.18 For monotherapy with nitazoxanide, another indirect inhibitor of HCV replication, data on viral load reduction are available only in patients infected with HCV genotype 4.19 However, indirect evidence for differences between HCV genotypes is available from combination therapies. Triple therapy with nitazoxanide, pegylated interferon, and ribavirin, after a lead-in phase with only nitazoxanide, resulted in different virologic response rates in patients infected with HCV genotype 1 compared with genotype 4 (5%-12% vs 64%, respectively).20, 21 Finally, silibinin, for which the mechanism of action is not yet entirely resolved, administered intravenously (20 mg/kg/day) for 7 days, led to a mean decline of the HCV-RNA concentration by 3 log10 IU/mL. So far, mainly genotype 1 and single patients with genotype 2, 3, and 4 infections have been investigated and no data on differences of antiviral activities for the different HCV genotypes are available.22
HCV subtypes and resistance
Differences in the HCV genome sequence are smaller between subtypes than between genotypes, resulting in smaller variations in antiviral activity of DAA agents between subtypes than genotypes.
However, differences between subtypes have been described in the activities of the linear protease inhibitors telaprevir and boceprevir. In clinical studies of telaprevir alone or in combination with pegylated interferon (with or without ribavirin), selection of resistant variants and viral breakthrough have been observed consistently more frequently in patients infected with HCV subtype 1a than subtype 1b.23, 24, 25 In the protease inhibition for viral evaluation (PROVE) 1 study (triple therapy: telaprevir, pegylated interferon alfa 2a, and ribavirin), 11 of the 12 patients who experienced a viral breakthrough were infected with HCV subtype 1a; only 1 of the 12 was infected with subtype 1b.25 This difference was shown to result from nucleotide differences at position 155 in HCV subtype 1a (aga, encodes R) vs 1b (cga, also encodes R). The mutation most frequently associated with resistance to telaprevir is R155K; changing R to K at position 155 requires 1 nucleotide change in HCV subtype 1a and 2 nucleotide changes in subtype 1b isolates (Figure 2). Selection of the resistant variant R155K to monotherapy with the NS3 inhibitor boceprevir also differed between patients infected with HCV subtypes 1a and 1b.26 Taken together, based on differences in codon usage for telaprevir and boceprevir several resistant variants were selected typically in subtype 1a- (V36M, R155K/T) or 1b- (R155Q, V170A) infected patients.23, 26 Differences between subtype 1a- and 1b-infected patients also were observed for macrocyclic NS3 protease inhibitors (ITMN191/R7227, BI201335).27, 28
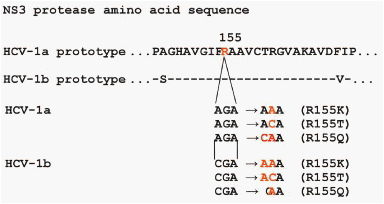
Figure 2. Molecular basis for different probabilities for selection of R155K/T/Q mutations in subtype 1a- vs subtype 1b-infected patients treated with a NS3 protease inhibitor. Selection of R155K/T requires 1 nucleotide exchange in subtype 1a but 2 nucleotide exchanges in subtype 1b patients. Selection of R155Q requires 1 nucleotide exchange in subtype 1b but 2 nucleotide exchanges in subtype 1a patients.
Nucleoside analogs that bind to the highly conserved, active site of NS5B seem to have an equal antiviral activity against different HCV subtypes; no differences have been reported for NM283, R1626, and R7128 in patients infected with HCV subtypes 1a or 1b.29, 30, 31
Polymerase inhibitors binding to less-conserved allosteric sites of NS5B have a greater risk of different antiviral activities and different resistance profiles in patients infected with different subtypes. Administration of BI207127, a nonnucleoside inhibitor of NS5B that binds to the thumb 1 site (NNI site 1), led to a viral decline in all patients infected with HCV genotype 1, without viral breakthrough during treatment for 5 days. However, BI207127 was reported to be less active against HCV subtype 1a than 1b.32 BILB1941, another thumb 1 site inhibitor, also had greater antiviral activity in patients infected with subtype 1b in comparison with subtype 1a.33
Filibuvir (PF-00868554) is a thumb 2 site (NNI site 2) inhibitor of NS5B. Differences in resistance to this drug were observed between patients infected with HCV subtypes 1a and 1b after 8 days of monotherapy.34 Also, triple therapy with filibuvir, pegylated interferon alfa 2a, and ribavirin resulted in a rapid virologic response more frequently in patients infected with subtype 1a (14 of 19) vs 1b (3 of 7) virus.35
Furthermore, differences in the reduction in median HCV-RNA levels have been reported between patients with HCV subtypes 1a (1.4-2.5 log10 IU) and 1b (2.6-3.2 log10 IU) who were treated for 3 days with the nonnucleoside inhibitor ANA598, which binds to the palm 1 site (NNI site 3).36 No viral breakthrough was detected during this short-term treatment but the observed differences of the antiviral activity could result from sequence differences between subtypes 1a and 1b in the ANA598 binding site. No host or HCV subtype-specific differences have been reported to affect the antiviral activity of HCV796, a palm 2 site inhibitor (NNI site 4) of NS5B. However, resistance data have been presented only from patients infected with HCV subtype 1a.37, 38 In vitro different resistant variants in subtype 1a and 1b isolates were selected during incubation with HCV-796.39
HCV isolates and resistance
Even within a given HCV subtype, variations in amino acid sequences are common and can affect the antiviral activity of an antiviral agent and the barrier of resistance.
For example, the pre-existing dominant strain in a patient's quasispecies can sometimes be an HCV variant with reduced sensitivity to NS3 inhibitors that bind to the active site of the protease. In different studies direct sequencing revealed resistant variants at positions V36, T54, R155, D168, and V170 of the NS3 protease in 0.2%-2.2% of patients as the pre-existing dominant strain.10, 40, 41, 42, 43 Patients with the R155K variant had a reduced response to telaprevir compared with patients without this variant and none of the 4 of 570 patients with this variant already present at baseline as the dominant strain achieved a sustained virologic response during treatment in the PROVE 1/2 studies.40 Studies using ultra-deep sequencing methods revealed that most amino acid variants that confer resistance to protease inhibitors are detectable within the HCV quasispecies already before treatment, but at low frequencies. Currently, it is unknown which frequency of a specific resistant variant influences effectiveness of treatment with an NS3 protease inhibitor.44
Mutations that confer in vitro resistance to nucleoside analogs that inhibit NS5B (S96T, S282T, N142T) were not detected in patients with HCV genotype 1 infections.10, 41, 45
Variants conferring resistance to nonnucleoside inhibitors that bind to different allosteric sites of the NS5B polymerase were detected in 0.6%-2.8% of patients by direct sequencing as pre-existing major variants.10, 34, 41, 45, 46 However, for nonnucleoside inhibitors resistance mutations also may occur as natural variants more frequently in patients infected with a specific subtype. For example, the C316Y mutation in NS5B is associated most frequently with resistance to HCV-796,37 although C316N also reduces sensitivity to HCV796 in vitro and this variant frequently is observed in many patients infected with HCV subtype 1b.45
Resistance mutations selected during treatment with DAA typically are associated with reduced replicative fitness. Interestingly, patients who have not yet received antiviral therapy but are infected with DAA-resistant variants as the pre-existing dominant strain have been reported to have high baseline concentrations of HCV RNA. Thus, it would be interesting to learn how the naturally occurring resistant variants achieved their high replicative capacities in the absence of drug selection.
Pharmacokinetics and Compliance
With the exception of silibinin all DAA agents are being developed for oral administration. Both the NS3 inhibitors that are in phase 3 studies, boceprevir and telaprevir, have a relatively short half-life, requiring a 3-times-per-day dosing interval.12, 52 Trough plasma levels were correlated with antiviral responses in phase 1 studies of these agents. Moreover, 6 of 10 patients who received 2 daily doses of 1250 mg telaprevir had viral breakthrough and resistance, compared with 1 of 8 patients who received 3 daily doses of 750 mg telaprevir (the subsequent standard therapy) in a phase 1 monotherapy study.23 Interim analysis after 12 weeks of triple therapy with telaprevir, pegylated interferon, and ribavirin showed no differences in the decrease of antiviral activity or viral breakthrough in patients treated 2 times daily with 1125 mg, compared with 3 times daily with 750 mg telaprevir. However, small numbers (39-42 patients) were enrolled in each cohort and differences might become visible only in larger studies, especially in nonresponders to previous pegylated interferon/ribavirin combination therapy.53
For patients with human immunodeficiency virus infection, compliance is one of the major factors that affect the development of resistance.54 Treatment failure from drug resistance has been decreased by reducing the administration of antiretroviral therapy from 3-times-daily to once-daily highly active antiretroviral therapy. Many DAA agents currently in phases 1 and 2 trials have a half-life that allows for once- or twice-daily dosing. Similar to human immunodeficiency virus treatment, ritonavir also is given to HCV patients along with several HCV NS3 inhibitors (ITMN191/R7227, SCH900518) to improve pharmacokinetic properties and prolong dosing intervals. However, about 10% of patients are intolerant to ritonavir because of its side effects. Thus, the addition of ritonavir to triple therapies of DAA, pegylated interferon, and ribavirin may enhance early discontinuation rates and thereby may reduce a patient's potential to achieve a sustained virologic response.
Combination Therapies
In human immunodeficiency virus and HBV infection combination therapies are used to overcome rapid selection of variants that are resistant to single agents and increase the odds of long-term suppression of viral replication. HCV variants with amino acid changes that confer resistance to DAA compounds retain their sensitivity to interferon alfa and non-cross-resistant drug classes in vitro.55, 56, 57
DAA plus pegylated interferon
Combination therapies of protease inhibitors with pegylated interferon alfa have been tested for their abilities to overcome resistance to single agents and for their additive or even synergistic antiviral activities. In a phase 1 study of 14 days of combination therapy with telaprevir and pegylated interferon alfa 2a, a continuous decline in HCV-RNA concentrations, without viral breakthrough, was observed. Furthermore, antiviral efficacy was at least additive, with a maximum decline of 5.5 log10 for the combination therapy, compared with 4.4 log10 for telaprevir monotherapy.58 Another study showed that combination therapy with boceprevir with pegylated interferon alfa 2b principally may be effective in patients who did not respond to previous interferon alfa-based treatment regimens.52 Combination therapies with pegylated interferon and other protease or polymerase inhibitors had similar effects. However, in the first phase 2 trial, which included a longer duration of therapy, up to 24% of patients who received combination therapy with telaprevir and pegylated interferon alfa 2a experienced a viral breakthrough with emergence of resistant HCV variants. In addition, a high relapse rate (48%) was observed.25, 59
Triple therapy DAA plus pegylated IFN plus ribavirin
The addition of ribavirin to combination therapy with a protease inhibitor and pegylated interferon alfa significantly increased antiviral activity, lowered relapse rates, and reduced viral breakthrough and selection for resistant variants. In a study that compared combination therapy with telaprevir and pegylated interferon alfa to triple therapy that included ribavirin, HCV-RNA levels were undetectable after 12 weeks in 62% and 80% of patients, respectively. Furthermore, viral breakthrough occurred during treatment in 24% of patients on double therapy and 1% on the triple therapy; virologic relapse occurred in 42% and 25% of the patients, respectively.59 Similar results were observed in other studies comparing dual vs triple therapies with the NS3 protease inhibitor boceprevir or the polymerase inhibitor R1626.30, 60, 61, 62 However, no studies have been conducted to test combination therapies with DAA and ribavirin but without interferon alfa. Future studies are needed to determine whether ribavirin exerts its antiviral activity only in the presence of interferon alfa or also if it can decrease viral load and prevent selection of resistant variants in combination with a DAA agent.
DAA combination therapies
Another possibility to suppress viral replication, avoid the selection of resistant variants and viral breakthrough, as well as potentially eradicate the virus are DAA combination regimens. Studies in the in vitro HCV replicon model and in chimpanzees have indicated the potential of combination therapies with protease and polymerase inhibitors. Indeed, using this approach, HCV was eradicated in a chimpanzee.57, 63 Results of the first clinical study of the combination of a polymerase inhibitor (R7128) and an NS3 inhibitor (R7227/ITMN191) were presented recently (INFORM-1 study). In this proof-of-principle study, patients were given both DAA agents for up to 2 weeks. HCV-RNA concentrations decreased by as much as 5.2 log10 IU/mL, no viral breakthrough was observed, and HCV RNA was undetectable by the end of the study in up to 63% of patients, measured using a highly sensitive assay.64 The potential of this combination therapy to suppress replication for longer durations, without viral breakthrough or selection of resistant variants, and to ultimately achieve a sustained virologic response, has to be shown in future studies.
Characterization of Resistant Variants Selected During Treatment With DAA
The resistance profile of an HCV quasispecies is best determined by analysis of the genome sequence that is targeted by the DAA agent using a highly sensitive clonal sequencing approach. By direct sequencing, only amino acid variations that occur in the HCV quasispecies at a frequency of approximately 25% can be detected, whereas cloning and sequencing of, for example, 80 isolates, viral variants present at approximately 5% of the population will be detected. In addition, the clonal approach also allows analysis of mutations linked on a given isolate. Moreover, because of the rapid viral turnover during HCV infection and because highly resistant variants typically are associated with a significantly impaired replicative fitness these variants will be overgrown by wild-type virus shortly after termination of antiviral therapy and/or compensated by additional mutations that enhance the replication efficiency. The likeliness of selection of isolates with compensated replicative fitness increases with the duration of DAA therapy without complete suppression of viral replication. Furthermore, for combination therapies resistant viral variants may be selected in the first days during treatment followed by a continuous viral decline because of a lack of cross-resistance between the different antiviral drugs.
It is not known whether resistant variants can persist in the long term as major or minor quasispecies in nonresponsive patients or those with viral breakthrough or relapse after DAA therapy. Thus, genotypic sequence analysis should be performed in the first days during therapy, directly after viral breakthrough, at the end of treatment, and during short-term and long-term follow-up evaluation. Initial characterization of the HCV resistance profile to DAA agents, using the highly sensitive clonal sequencing method, is mandatory. However, other methods for sequence analysis are available (ie, direct sequencing, TaqMAMA, hybridization assays, restriction enzyme assays, deep sequencing methods). It remains to be established which approach will be most useful to identify DAA agent resistance mutations in clinical practice.65
The first characterization of HCV resistance variants selected during monotherapy with telaprevir and identified by the clonal method identified mutations that were not predicted with the HCV replicon assays.23, 66 Previous in vitro replicon studies identified the A156S/V/T mutation in NS3, but not mutations at V36, T54, or R155, which were selected in patients treated with telaprevir. High doses of telaprevir were used in the in vitro replicon studies, whereas lower concentrations of boceprevir selected for the T54A and V170A mutations in NS3 in the HCV replicon system. Mutations at position Q41 and F43 of NS3 were identified in the replicon assay, but these variants were not selected in patients treated with telaprevir or boceprevir so far.67 Mutations in NS3 that conferred resistance to boceprevir, such as V55A, also were not predicted from in vitro studies but were detected using highly sensitive genotypic analysis of the HCV quasispecies in patients from the phase 1 monotherapy study.26
Interestingly, for both telaprevir and boceprevir during monotherapy a large number of mutational variants were selected (n = 25-31) and restrictions (increase of more than 5% frequency from baseline, detection in more than 1 patient, decline after the end of treatment) were made for further analysis.23, 26, 68 Finally, mutations at 6 amino acid positions (V36, T54, V55, R155, A156, and V170) were associated clearly with resistance to these protease inhibitors. A potential importance of the multiple additional low-frequency variants at other positions has to be investigated in future studies.
After genotypic characterization, levels of resistance to the antiviral agent used in the clinical study, and potential cross-resistance to other direct antivirals must be determined by phenotypic assays. Different replicon- and enzyme-based phenotypic assays have been used to determine the IC50/EC50 values of DAA agents for HCV replication or enzymatic functions of wild-type and mutant HCV proteins.23, 26, 27, 69, 70 The majority of replicon assays is based on the HCV con1b isolate with restrictions for variants detected in subtype 1a isolates and other genotypes.39 Furthermore, typically only the detected variant, alone or in combination with another resistant variant, rather than the entire NS3 protease gene from a given patient, is introduced into NS3 for analysis in a phenotypic assay.
The data from clonal genotypic and phenotypic resistance analyses of telaprevir and boceprevir from phase 3 trials have been published.23, 24, 26 However, the resistance profiles of the majority of DAA agents have not been characterized fully. We summarize the available resistance genotypic and phenotypic resistance profile of DAA agents that are in phase 1, 2, or 3 trials.
NS3 Protease Inhibitors
BILN2061 (Ciluprevir) was the first NS3 protease inhibitor investigated in clinical studies. BILN2061 showed a high antiviral activity in genotype 1 and a variable antiviral activity in genotype 2/3-infected patients after dosing for 2 days. However, further development of BILN2061 was stopped and no clinical resistance data are available.71, 72, 73
Telaprevir
Telaprevir has a high antiviral activity (Figure 1A) but a medium genetic barrier to resistance.12 During monotherapy of genotype-1-infected patients with different doses of telaprevir for 14 days mainly viral variants at 6 positions within the NS3 protease were selected rapidly (Figure 3).23 These variants were detected either alone or in combination on the same isolate (mainly positions 36+155 or 36+156). Phenotypic analysis showed low- to medium-level (V36, T54, R155, A156S) and high-level (A156V/T) resistant variants. Molecular-based mechanisms of resistance have been proposed.66, 74, 75, 76 In more than 80% of the isolates of the HCV quasispecies at the end of treatment resistance mutations were detected. High-level resistant variants typically were observed in patients with higher telaprevir exposure and more pronounced viral decline. Generally, an inverse correlation between the level of telaprevir resistance conferred by a single mutation and replicative viral fitness was observed. Double mutants (V36A/M + R155K/T) seem to have a compensatory effect with an increase of replicative fitness together with a relatively high level of resistance.23, 24 After termination of treatment with telaprevir the number of wild-type isolates rapidly increased within the first 7-10 days of follow-up evaluation. After 3-7 months no high-level resistance was observed anymore and the majority of resistant variants were replaced by wild-type virus.23 However, in single patients V36 and R155 variants still were detectable at low levels and in a recent report on long-term follow-up evaluation even after up to 3 years in single patients low- to medium-levels of V36 and R155 variants were observed.77
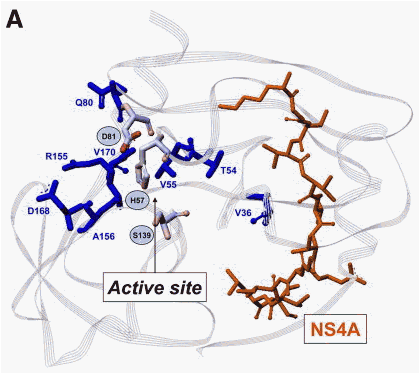
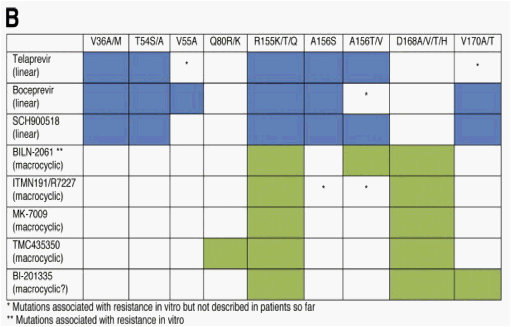
Figure 3. Amino acid positions within the NS3/4A protease associated with (A) resistance mutations to different NS3 protease inhibitors and a (B) cross-resistance table of different NS3 protease inhibitors based on mutations selected in patients from clinical studies and/or from in vitro studies. Amino acids forming the catalytic triad (H57, D81, S139) are highlighted. *Mutations associated with resistance in vitro but were not described in patients. **Mutations associated with resistance in vitro. Resistance mutations of linear NS3 protease inhibitors are shown in blue and resistance mutations described for macrocyclic NS3 protease inhibitors are shown in green.
During combination therapy of telaprevir with pegylated interferon alfa A156T/V variants were detectable in the initial viral decline phase but viral levels continued to decline.24 During triple therapy with telaprevir, pegylated interferon alfa 2a, and ribavirin in 3% of the patients (5 of 163) a viral breakthrough with wild-type virus in 1 patient but high-level resistant variants in 4 patients (V36M+R155K, A156T) was observed. Furthermore, in patients with relapse after the end of treatment also low- and high-level resistant variants with R155K as the main resistance mutation were detected.59
Generally, the likeliness of a viral breakthrough is correlated with the response to pegylated interferon alfa/ribavirin combination therapy. The majority of patients with a virologic null-response (<1 log10 decline at week 4 or <2 log10 decline at week 12) to pegylated interferon alfa/ribavirin combination therapy who were retreated with triple therapy including telaprevir showed a viral breakthrough. In partial nonresponders (>2 log10 decline at week 12 but HCV-RNA positive at week 24) only several viral breakthroughs were observed during triple therapy and in patients with a prior relapse after standard combination therapy no viral breakthrough was observed during re-treatment with triple therapy including telaprevir.78 Thus, for patients with null-response to pegylated interferon alfa/ribavirin, triple therapy is a form of functional monotherapy with the direct antiviral drug and this is associated with a high probability of viral breakthrough. Future studies have to investigate the possibility to select the optimal patients for triple therapy perhaps with a lead-in phase to characterize the responsiveness to pegylated interferon alfa/ribavirin alone or together with other response predictors (baseline viral load, IL28 polymorphism. This would on the one hand enable the possibility to avoid viral breakthrough with selection of resistant variants and perhaps on the other hand for patients with favorable response predictors no need of the addition of a DAA or very short triple therapies.
Telaprevir was investigated also in genotype 2-, 3-, and 4-infected patients with lower (genotype 2) or virtually absent (genotype 3, 4) antiviral activities.13, 14 Data on resistant variants selected in patients infected with genotype 2, 3, and 4 isolates will be presented at an upcoming meeting.
Boceprevir
Boceprevir has a medium antiviral activity (Figure 1A) and a medium genetic barrier to resistance.79 During monotherapy with 400 mg boceprevir 2 or 3 times daily for 14 days, mainly viral variants at 6 positions within the NS3 protease conferring low- to medium-level resistance were observed (Figure 3). As for telaprevir and also for boceprevir an increasing number of selected variants correlated with a higher viral decline. Interestingly, the mean frequency of wild-type variants was still 86% at the end of treatment and the high-level resistant variant A156T was not observed.26 The currently used dose of boceprevir in the phase 3 trials is 800 mg 3 times daily. However, no monotherapy data with this dose are available and it is unknown whether with the higher dose also high-level resistant variants will be selected.
Genotypic analyses show a largely overlapping cross-resistance profile of boceprevir and telaprevir (Figure 3). However, for example, the V170A variant conferred a higher level of resistance to boceprevir in comparison with telaprevir and this variant was not described to be a frequent variant in patients who received telaprevir.23, 24, 26 Shortly after the end of boceprevir treatment the frequency of wild-type variants increased to 96%. However, in a number of patients 1 year after the end of boceprevir dosing and in single patients even 4 years after the end of boceprevir dosing, either alone or in combination with pegylated interferon, resistant variants still were detected in the HCV quasispecies by clonal sequence analysis.80 Boceprevir also was investigated in genotype 2- and 3-infected patients. However, antiviral activity was reduced in these genotypes and no resistance analysis is available from these studies.
Triple therapy with boceprevir, pegylated interferon alfa 2b, and ribavirin in a large phase 2 study in genotype 1 treatment-naive patients for 2848 weeks was associated with viral breakthrough and selection of resistant variants in 7%-12% of the patients.61 Patients who received a lead-in treatment with pegylated interferon and ribavirin alone for 4 weeks before the addition of boceprevir for another 24 or 44 weeks showed lower viral breakthrough rates (4%-5%).61 However, to answer the question whether a lead-in phase with peginterferon and ribavirin alone will generally reduce the probability of a viral breakthrough, analysis of the phase 2 and 3 studies with boceprevir, telaprevir, and other DAA have to be awaited.
Other NS3 protease inhibitors (R7227ITMN191, MK7009, BI201335, TMC435350, SCH900518, BMS-650032, PHX1766)
Several NS3 protease inhibitors currently in phase 1/2 development showed high antiviral activities in monotherapy studies (Figure 1A)81, 82, 83, 84, 85, 86, 87 and initial resistance analyses have been presented at scientific meetings.27, 28, 84, 85, 88, 89 Taken together, differences between linear tetrapeptide and macrocyclic inhibitors binding to the active site of the NS3 protease have been revealed (Figure 3). However, R155 is the main overlapping position for resistance, and different mutations at this amino acid site within the NS3 protease confer resistance to all protease inhibitors that are currently in advanced clinical development (Figure 3).
Polymerase Inhibitors
Several nucleoside inhibitors targeting the active site of the HCV-RNA-dependent RNA-polymerase as well as a number of nonnucleoside inhibitors binding to different allosteric sites of the NS5B polymerase are currently in clinical development (Table 1).
Nucleoside analogs
Valopicitabine (NM283, 2'-C-methylcytidine/NM107), the first nucleoside inhibitor investigated in patients with chronic hepatitis C, showed a low antiviral activity (Figure 1B).29 Because of gastrointestinal side effects the clinical development of NM283 was stopped. Very sparse clinical data on resistance are available but from in vitro assays S282T mutation in the NS5B polymerase is known to confer resistance to NM283 (Figure 4).90
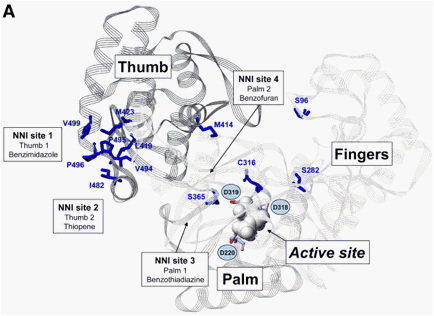
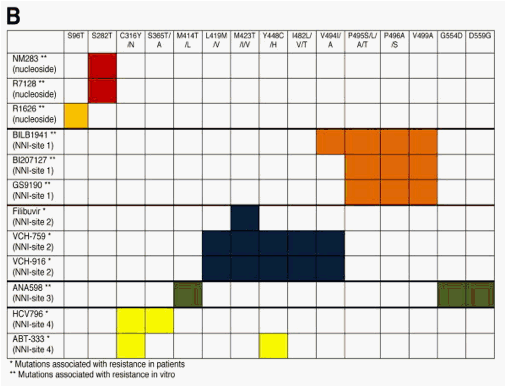
Figure 4. Amino acid positions within the NS5B polymerase associated with (A) resistance mutations to different nucleoside and nonnucleoside inhibitors and a (B) cross-resistance table of different NS5B polymerase inhibitors based on mutations selected in patients or in vitro. Amino acids forming the catalytic triad (D318, D319, D220) are highlighted. *Mutations associated with resistance in patients. **Mutations associated with resistance in vitro. Mutations of 2'-C-methylcitidine and 4'-azidocytidine nucleoside inhibitors are shown in red and pink, respectively. For NNI-site 1, 2, 3, and 4 inhibitors resistance mutations are shown in orange, dark blue, dark green, and yellow, respectively.
The second nucleoside inhibitor to be reported in patients with chronic hepatitis C was R1626 (4'-azidocytidine/PSI-6130). A phase 1 study in genotype 1-infected patients observed a high antiviral activity at high doses of R1626 in genotype 1-infected patients (Figure 1B).30 No viral breakthrough with selection of resistant variants was reported from monotherapy or combination studies with pegylated interferon ± ribavirin.30, 62 In HCV replicon studies S96T mutation within the NS5B polymerase conferred resistance to R1626.90 Because of severe lymphopenia and infectious disease adverse events further development of R1626 was stopped.
R7128, another nucleoside polymerase inhibitor with antiviral activity in genotype 1-infected patients in phase 1 monotherapy studies,91 currently is being investigated in phase 2 clinical trials in genotype 1-, 2-, and 3-infected patients in combination with pegylated interferon and ribavirin as well as in combination with the protease inhibitor R7227/ITMN191. Also from these studies no viral breakthrough with selection of resistant variants was reported. In vitro, different resistant variants for R7128/NM283 (S282T) and R1626 (S96T) have been selected and cross-resistance was excluded (Figure 4).90
Recently, 2 other polymerase inhibitors (PSI-7851 and IDX184) were evaluated in phase 1 clinical trials in patients with chronic hepatitis C (Figure 1B). However, no data on resistance have been presented for these drugs so far.92, 93
Nonnucleoside analogs
At least 4 different allosteric binding sites have been identified for inhibition of the NS5B polymerase by nonnucleoside inhibitors (Table 1). A number of mutations conferring resistance to the different chemical classes of nonnucleoside inhibitors have been observed mainly in in vitro studies (Figure 4).
NNI-site 1 inhibitors (thumb 1/benzimidazole site)
BILB1941, BI207127, and MK-3281 are NNI-site 1 inhibitors that have been investigated in clinical phase 1 trials with low to medium antiviral activities (Figure 1C).32, 33, 94, 95 No viral breakthrough with selection of resistant variants has been reported during 5 days of treatment with BILB1941 or BI207127.32, 46 However, preexisting NS5B substitutions known to reduce sensitivity were identified for BILB1941 (V494I/A, I424V, P496A) and resistance mutations of NNI-site 1 inhibitors were characterized in vitro (Figure 4).96, 97
NNI-site 2 inhibitors (thumb 2/thiophene site)
Filibuvir (PF-00868554) is a NNI-site 2 inhibitor with medium antiviral activity in a phase 1 study (Figure 1C).98 Pre-existing variants conferring reduced sensitivity to filibuvir were observed in single patients (M426A). Several filibuvir resistant variants have been described in HCV replicon studies.99 During treatment a high number of different variants was observed. However, variants at position M423 known to confer resistance to filibuvir from in vitro studies were selected in the majority of patients (Figure 4).34, 99 In a subsequent triple-therapy trial with filibuvir, pegylated interferon alfa 2a, and ribavirin for 4 weeks, a viral breakthrough was observed in 5 of 26 patients.35
VCH-759, VCH-916, and VCH-222 are 3 other NNI-site 2 inhibitors with antiviral activities in monotherapy studies (Figure 1C).16, 100, 101 For VCH-759 as well as VCH-916 viral breakthroughs with selection of resistant variants conferring high (M423T/V/I) and medium (L419V/M, I482L/V/T, V494A/I) levels of resistance were observed (Figure 4).16, 102 In the majority of patients who experienced a viral breakthrough resistance mutations were found to remain present up to 13 days despite a high HCV replication rate and a reduced replication fitness of the variants in vitro.16
NNI-site 3 inhibitors (palm 1/benzothiadiazine site)
ANA598 is a NNI-site 3 inhibitor that displayed antiviral activity during treatment of genotype 1-infected patients (Figure 1C).36 No viral breakthrough was observed during this short-term monotherapy trial. However, several variants were described from in vitro replicon studies to confer resistance to ANA598 (Figure 4).103
NNI-site 4 inhibitors (palm 2/benzofuran site)
Monotherapy with the NNI-site 4 inhibitor HCV-796 showed low antiviral activity in genotype 1-infected patients (Figure 1C).38 Viral breakthrough was associated with selection of resistant variants that conferred medium to high levels of resistance in phenotypic analysis37, 104 (Figure 4). For GS-9190 low antiviral activity was observed in a clinical study and variants conferring resistance were identified in the β-hairpin of the polymerase.94, 105 ABT-333, another palm site inhibitor, showed antiviral activity in patients with genotype 1 infection (Figure 1C) and from in vitro replicon studies C316Y and Y448C were observed as main resistance mutations.106, 107 In addition, S556G was reported to be associated with viral breakthrough in a clinical study (Figure 4).106
NS5A Inhibitor
In a single ascending dose study it was shown that inhibition of the NS5A protein with BMS-790052 leads to a sharp initial decline of HCV-RNA concentrations (Figure 1D).108 BMS-790052 is the first NS5A inhibitor binding to domain I of the NS5A protein, which was shown to be important for regulation of HCV replication.109 No clinical data on resistance to this class of drugs have been presented yet and results of multiple dose studies have to be awaited.
Cyclophilin Inhibitors
Cyclophilins are functional regulators of the HCV NS5B polymerase.110 Debio-025, NIM811, and SCY-635 are cyclophilin A inhibitors, which block HCV replication in vitro.50, 111, 112 For Debio-025 antiviral activities in patients infected with different HCV genotypes (1-4) was shown during monotherapy and in combination studies with pegylated interferon (Figure 1D).18, 113 No viral breakthrough was observed in these studies. However, selection of HCV variants with mutations clustering in the NS5A gene resistant to Debio-025 was possible in the HCV replicon system.51, 112 Another cyclophilin inhibitor SCY-635 also showed antiviral activity in genotype 1-infected patients (Figure 1D).50 During this 14-day monotherapy study no viral breakthrough was reported. In vitro mutations within the NS5A (T17A, E295K, V444A) and NS5B (T77K, I432V) protein conferred resistance to SCY-635.114 No viral decline in genotype 1-infected patients treated with NIM811 alone was observed in a phase 1 study (Figure 1D).115 However, selection of cell lines conferring resistance to NIM811 without disclosure of the mutations observed was shown in the replicon system.111 Taken together, cyclophilin inhibitors may have the potential of broad antiviral activity to different HCV genotypes. However, selection of resistant variants is possible and the natural- and treatment-induced frequency of mutations conferring resistance to the different cyclophilin inhibitors have to be explored.116, 117
Nitazoxanide
Nitazoxanide is a thiazolide anti-infective with activity against anaerobic bacteria, protozoa, and viruses. It is approved for treatment of cryptosporidiosis and lambliasis in the United States. In vitro nitazoxanide was shown to inhibit HCV replication and the mechanism of action was proposed to be induction of double-strand RNA activated protein kinase.118 In a placebo-controlled study monotherapy with nitazoxanide for 24 weeks led to a mean reduction of HCV-RNA viral load in genotype 4-infected patients by 1.55 log10 IU/mL with 30% of patients having undetectable HCV RNA at the end of treatment.19 Studies in genotype 1-infected patients with a lead-in monotherapy with nitazoxanide for 4 weeks have been performed. However, no virologic response data in genotype 1-infected patients during monotherapy with nitazoxanide have been reported (Figure 1D).21, 119 Patients experiencing viral breakthroughs were not reported from monotherapy studies. However, in vitro studies suggested the potential of resistance to nitazoxanide most likely was conferred by changes in host proteins and not by mutations in the virus.118
Silibinin
The mechanism of action of silibinin is unknown but inhibition of NS5B polymerase as well as antiviral and anti-inflammatory activities shown in NS5B polymerase assays, in human peripheral blood mononuclear cells, and the HCV replicon, respectively, have been reported.120, 121 Monotherapy studies with daily intravenous application of silibinin in genotype 1-, 2-, and 3-infected patients for up to 14 days showed good antiviral activities (Figure 1D).22 Studies in larger cohorts of patients including resistance analysis are underway.
Conclusions
The development of DAA agents has increased the number of therapeutic options for patients with HCV infection. Data from phase 2 studies have shown that rates of sustained virologic response significantly improve in patients given triple therapies of a DAA agent, pegylated interferon, and ribavirin. However, these approaches also have many restrictions and raise some new problems and questions. Because of the high level of genomic heterogeneity among HCV subtypes and isolates, DAA agents will not have similar levels of activities in all patients. Most reagents are being developed to treat patients infected with HCV genotype 1. Although sustained virologic response rates are increasing among these patients, triple therapy is not effective in all patients infected with genotype 1 because of side effects, nonresponsiveness to pegylated interferon or ribavirin, or development of resistance. It is important to increase our understanding of mechanisms of resistance mutations, viral breakthrough, and relapse after therapy with DAA agents. Improving our knowledge of these processes will require more precise and earlier characterization of resistance profiles of patients given monotherapy, long-term follow-up data, and studies with combination therapies. Studies also are underway to test the effects of combinations of DAA agents without interferon alfa or ribavirinčthese trials will provide valuable information about the ability of DAA agents to suppress HCV replication without development of resistance or even to eradicate the virus.
|
|
| |
| |
|
|
|